
Grantee Research Project Results
2006 Progress Report: Species-Specific Xenobiotic Metabolism Mediated by the Steroid and Xenobiotic Receptor SXR
EPA Grant Number: CR830686Title: Species-Specific Xenobiotic Metabolism Mediated by the Steroid and Xenobiotic Receptor SXR
Investigators: Blumberg, Bruce
Institution: University of California - Irvine
EPA Project Officer: Hahn, Intaek
Project Period: January 1, 2003 through December 31, 2005 (Extended to December 31, 2007)
Project Period Covered by this Report: January 1, 2005 through December 31,2006
Project Amount: $949,986
RFA: Issues in Human Health Risk Assessment (2001) RFA Text | Recipients Lists
Research Category: Human Health
Objective:
The overall aim of the proposed research is to aid in providing a molecular basis for understanding the commonalities and differences in how humans and model animals respond to chemical exposure. We hypothesize that activation of the nuclear steroid and xenobiotic receptor (SXR)/pregnane xenobiotic receptor (PXR), and consequent effects on metabolism, is the mechanism underlying the differential susceptibility of humans and laboratory animals to environmental chemicals. The specific objectives of the proposed research are to: (1) characterize the commonalities and differences in the response of human and rodent SXR/PXR; (2) identify functional differences in the activation and/or regulation of SXR/PXR among humans and between commonly used strains of laboratory mice; (3) determine whether the compounds are metabolized in vivo; and (4) identify target genes regulated by SXR/PXR as a response to environmental chemical exposure.
Progress Summary:
During this no-cost extension year, we continued to make excellent progress on aims 1, 2, and 4 above. Two peer-reviewed reviews on endocrine disrupting chemicals (EDC) were published in Molecular Endocrinology (Tabb and Blumberg, 2006) and Endocrinology (Grun and Blumberg, 2006). Two manuscripts from our laboratory were published in the high profile journals Molecular Endocrinology (Grun, et al., 2006) and The Journal of Clinical Investigation (Zhou, et al., 2006b). Three other collaborative studies were facilitated by this award and also appeared in 2006 (Zhou, et al., 2006a; Gunderson, et al., 2006; Zhou et al., 2006c). Continuing progress on the aims listed above is described below.
Identification of SXR Homologs from Other Species
We completed testing of the SXR “zoo” against a panel of xenobiotic compounds. The set of compounds tested was expanded from the 17 shown in the previous progress report to 27, which is the final total. The receptors tested included human, mouse, rat, rabbit, beagle, marmoset, Japanese monkey, crab eating monkey, chicken, Japanese quail, Xenopus, carp, fathead minnow, fugu, zebrafish and medaka. The results are summarized in Table 1. To our knowledge, these results represent the largest study of the effects of xenobiotic chemicals on any receptor system ever reported. Moreover, the number of species included is unprecedented in the nuclear receptor field. SXR homologs from 17 species were tested, including common toxicological models such as beagle dog, rat, fathead minnow, and carp as well as laboratory models such as mouse, chicken, quail, zebrafish, and medaka. In addition, a group of monkeys was studied (marmoset, Japanese monkey, crab eating monkey), which significantly expands the data available for primate SXRs. Each of these receptors was tested against a panel of 27 chemicals of considerable interest at three different doses, in triplicate and replicated in three experiments. The compounds tested include an extensive series of phthalates (8), pesticides, herbicides and fungicides (16), plastics components other than phthalates (4), and other miscellaneous compounds of interest (2). The fish receptors continue to be incredibly diverse from each other. Fathead minnow and carp are very much closer to each other than to other fishes and are quite different from zebrafish, fugu, and medaka. Notably, fathead minnow and carp are somewhat similar to the other vertebrate models, including human, whereas zebrafish, fugu, and medaka are not. This suggests that despite their utility as genetic model organisms, zebrafish and medaka are wholly unsuitable for toxicological studies if the results are intended to be extrapolated to humans and other mammals. Based on our results, it can be argued that the fish are not good models for each other, let alone models for effects on higher vertebrates and humans.
Table 1. Activation of the SXR “Zoo” in Transient Transfection Assays. CMX-GAL4-LBD constructs were transiently transfected into COS-7 cells and treated with ligands at the indicated concentrations. Data are expressed as fold activation over solvent controls and represent the mean of triplicate experiments. The results are color coded. Red indicates that the compound was toxic at the tested concentration, yellow indicates activation below the level we consider significant (< 2 fold induction), gray compounds are weakly active (2–4 fold induction), blue compounds are moderately active (4–10 fold induction), green compounds are highly active (> 10 fold induction). The bottom five rows, rifampicin (RIF), clotrimazole, PCN, 5β-3,20 pregnane dione, and p-hydroxy benzoic acid butyl ester are positive controls for activation of receptors in the columns where they are used.
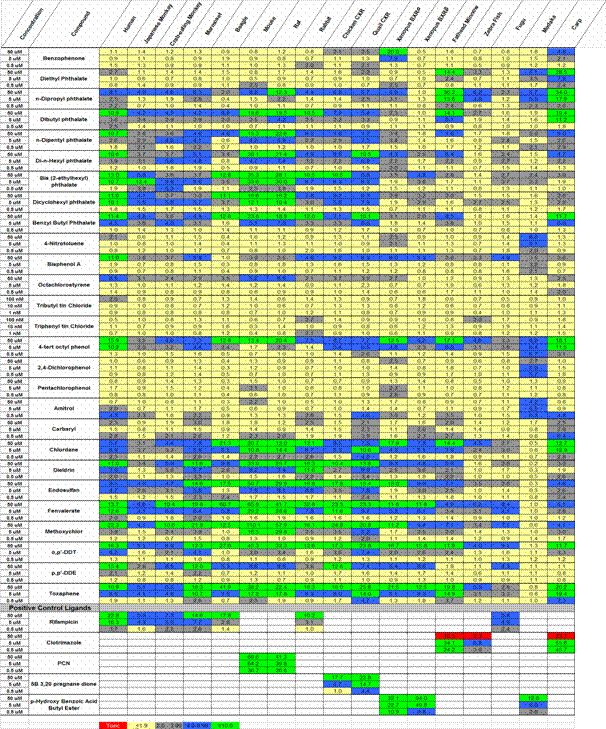
These studies are now complete and the manuscript is in preparation for Environmental Health Perspectives. We anticipate submission early in 2007, and expect that the review and publication process will be complete in 2007.
Identification of SNPs in Human and Mouse SXR
Our characterization of various mouse strains and human cell lines for single nucleotide polymorphisms (SNPs) in SXR/PXR showed that there were no differences in the coding sequences, suggesting that differences in the response of SXR/PXR to ligands is not responsible for observed differences in xenobiotic metabolism. In contrast, 14 SNPs have been identified that alter the protein coding region of human SXR. We have reconstructed all of these SNPs into the full-length receptors and are testing their response to a subset of the compounds shown in Table 1 (largely those that strongly activate wild type SXR). These studies are still in progress but preliminary results show that two of the SNPs are of particular interest for the study of EDC and human risk assessment.
Published reports have tested a few of the known SNPs on a small panel of SXR activators and concluded that the differences in response are relatively modest. In contrast, our initial results show that two of the SNPs previously characterized as having modest effects on transactivation, D163G and A370T, actually have rather drastic effects on activity. D163G is nearly inactive, being 200 fold less sensitive to ligands than the wild type receptor: EC50 = 2130 μM versus 10.8 μM for wild type in response to RIF. In contrast, A370T is nearly twice as active as wild type (Figure 1).
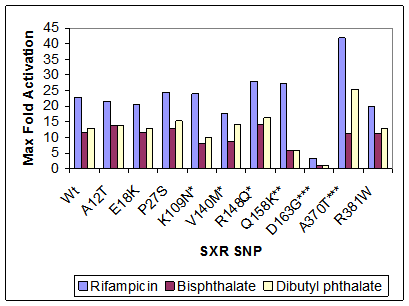
Figure 1. Maximum Fold Activation of Wild Type and SXR SNPs in Response to Three Ligands. *significant for one ligand, **significant for two ligands, ***significant for three ligands.
To evaluate the overall effect of the SNPs on transcriptional activity, we can broadly group them into four categories based on maximum fold-induction and EC50 values.
- Category 1 includes the SNPs that had maximum activations as well as EC50 values that did not differ significantly from wild type (p > 0.05), that is, those that show a relatively normal response. A12T, E18K, P27S, K109N, R148Q, and R381W all do not behave significantly differently from wild type when tested with RIF.
- Category 2 contains the polymorphisms that have similar maximum activation but significantly different EC50 values, making them more sensitive to ligand. Only V140M and Q158K fall into this group. Since a lower dose of RIF is required to achieve the same response, we infer that these mutations have increased the sensitivity of the receptor to ligand treatment.
- Category 3 includes polymorphisms that had different maximum activation but similar EC50 values. A370T is the only SNP in this group. Since the receptor reaches half-maximal activation at the same RIF dose, but has a higher overall magnitude of activation, we conclude that the SNP enhances receptor activity without affecting ligand binding.
- Category 4 includes SNPs that significantly differ from wild type, both in maximum activation and EC50 values. Only D163G fell into this category, and it was less sensitive to ligand and less well activated. This is consistent with its performance in response to all three ligands applied.
About half of the SNPs (A12T, E18K, P27S, and R381W) showed no significant change in response to bisphthalate. The others (K109N, V140M, R148Q, Q158K, and A370T) show a significant change in maximum activation, with decreased activation for all but A370T, but no significant difference with respect to EC50 values. Dibutyl phthalate elicited the least number of differences compared with wild type SXR. Seven of the 10 SNPs showed no significant difference in activation or EC50. Q158K showed a reduction in activation, whereas A370T exhibited a higher degree of activation, but neither SNP demonstrated a change in sensitivity. This result has been borne out when testing the entire panel of 14 SNPs.
Due to the major role played by SXR in maintaining chemical homeostasis in the body, our findings may have considerable relevance, both clinically and for human risk assessment. For example, individuals carrying the D163G or A370T SNPs could face greater incidence of drug interactions including reduced drug clearance or loss of therapeutic efficiency, which are important concerns. At the same time, they would be predicted to have altered sensitivity to xenobiotic exposure. A370T carriers would be much less sensitive to compounds that are detoxified by an SXR-dependent pathway and correspondingly more sensitive to compounds rendered more toxic in response to SXR activation. Exposure to SXR activators could also lead to the increased metabolism of endogenous steroids, thereby disrupting the body’s homeostatic mechanism. Carriers of D163G should display the opposite effect. In these individuals, SXR function would be severely compromised. Therefore, severe toxic reactions to drugs and xenobiotics could occur as a result of compromised clearance resulting from the reduced function of this SXR. Further investigation involving more ligands and SNPs will illuminate the effects of mutations in SXR on human health. The completion of these studies, now in progress, will be very important in understanding the spectrum of human responses to chemicals mediated through SXR. We anticipate that data collection will be complete at the end of January 2007, and a paper will be ready to be submitted in mid 2007.
A second set of investigations underway concerns the effects of the SXR SNPs on the activity of NF-κB. The mutually negative interactions between these pathways have broad relevance for toxicology, risk assessment, and clinical pharmacology (Zhou, et al., 2006b). One obvious question is to ask whether SXR SNPs have any effect on the repression of NF-κB signaling. This is likely to be an unappreciated mechanism through which individuals differ in their response to chemicals. The full-length SXR constructs are being transfected into HepG2 (liver) cells, and the effects on the activation of an NF-κB reporter in response to TNF-α evaluated. Our hypothesis is that one or more of the SNPs will alter the ability of SXR to negatively cross-talk with NF-κB. We are also testing mutants in SUMOylation sites in SXR as potential mediators of altered interaction with NF-κB. These studies are underway in parallel to the xenobiotic activation studies reported above. Both are expected to be completed early in 2007 and the manuscripts submitted in mid 2007.
Computational Modeling of SXR Ligands
Our original proposal was to generate a computational model for SXR that would enable the a priori prediction of whether a compound would be likely to activate or antagonize SXR. We succeeded in a limited way with polychlorinated biphenyls (PCBs) and this was published previously. Our efforts to generalize the model were frustrated by the reluctance of our collaborators to utilize the quantitative structure-activity relationship (QSAR) model they developed to predict the activity of compounds outside the chemical space of the training set on SXR function. This rendered the model ineffective for the intended use. Over the past year, we have established a collaboration with Professor Pierre Baldi and his colleagues at University of California at Irvine toward this end. Professor Baldi is a bioinformatician whose laboratory is expert in the area of computational modeling, among other things. The Baldi group has created a computational model for SXR binding that appears to be able to discriminate SXR agonists and antagonists. They have created a database of more than 5 million commercially available compounds (ChemDB; http://cdb.ics.uci.edu/CHEMDB/Web/index.htm Exit ) and docked a large subset of these into the SXR ligand binding domain and predicted which will be high affinity ligands. Using the data we previously generated from PCBs and a variety of other compounds, a model was developed that predicted which compounds could interact with SXR and which ones might be agonists versus which were likely to be antagonists. This model was tested by analyzing all 209 PCBs. It predicted that PCBs 69, 115, 116, 129, 185, 194, and 205 (which were not tested or identified in our previous experiments) would bind to SXR. Of these, PCBs 184, 194, and 205 were predicted to be antagonists. The others were predicted to bind, but the model did not identify which might be agonists or antagonists. We tested these predictions experimentally and observed that 184, 194, and 205 are, in fact, antagonists and that the others are agonists. These results provide an important validation of the computational model and enable its refinement. We are currently testing another group of predicted ligands from the National Cancer Institute (NCI) database to further test and refine the model. These experiments are also expected to be completed by mid 2007 and a paper submitted before the end of the year.
Identification of SXR Target Genes
We have collected microarray datasets from hepatocytes of five subjects treated with RIF. We are continuing our analysis of these target genes to characterize the bona fide targets. The study has also been expanded to include the computational modeling. Dr. Baldi and his colleagues have used their SXR binding model and performed docking with all of the cellular metabolites listed in the Kyoto Encyclopedia of Genes and Genomes (KEGG metabolites). We have cross-referenced this list with genes in KEGG pathways up- and down-regulated by SXR activators and identified six candidates for further study. These may reveal new cellular pathways in which the metabolites regulate SXR to modulate the expression of key enzymes in the pathway. This would be an extremely exciting result and a novel use of microarray and chemoinformatic techniques. It will also facilitate the publication of our microarray data in a higher profile journal than might otherwise be possible.
Characterization of the “Fully-Humanized” SXR Mouse
As mentioned in the previous report, we collaborated with Dr. Jun Kanno and his colleagues at the National Institutes of Health Sciences in Tokyo, Japan to generate a mouse that is predicted to show a “fully-humanized” response to human SXR activators. This (hSXRki) animal showed a humanized response to RIF in the liver and intestine. It is generally accepted that eight generations of back crossing are required to fully stabilize a strain. Our colleagues have been back crossing these mice, which are now at the eighth generation. The colony is being expanded and testing is underway. We anticipate that a manuscript will be submitted in the first half of 2007 describing two experiments that are key for our understanding of human risk assessment. The first set of experiments characterizes the response of these mice to ligands and will establish that they are excellent models for human exposure to chemicals. As noted above, preliminary studies show that these mice have a humanized response to RIF. This is a key point, because there are no other such models available. The best alternative is a transgenic mouse expressing human SXR in the liver and intestine produced by Dr. Wen Xie and his colleagues at the University of Pittsburgh. These mice are reported to show a great deal of individual variation in their response to RIF (Dave Eaton, personal communication). Moreover, they do not express SXR under the control of its normal promoter but rather as a transgene under the control of a promoter expressed in the liver and small intestine. This limits the utility of the mouse as a general model.
The second set of experiments tests the effect of the soy phytoestrogen genistein. There are abundant data from rodents to suggest that exposure to soy phytoestrogens has deleterious pro-estrogenic effects, including increased incidence of reproductive tumors. In contrast, human epidemiological data suggest that increased consumption of soy phytoestrogens is neutral or beneficial. Genistein, the major soy phytoestrogen, is a human-selective SXR activator which may result in its differential metabolism in rodents versus humans. Using the humanized mouse will enable us to definitively test whether differences in the ability of genistein to activate SXR explains the controversial observations in rodents versus humans. Dose response experiments are currently being conducted by Dr. Kanno and his colleagues, and the entire suite of experiments is expected to be concluded in early 2007. The use of this model will enable us to address Objective 3—whether or not compounds are metabolized in vivo in response to activation of SXR. Such experiments have been long delayed waiting for these mice and are now feasible.
Mutually Inhibitory Interactions Between SXR and NF-κB Signaling Pathways
As described in the previous progress report, we discovered a novel and unexpected interaction between SXR and NF-κB signaling pathways. These results have now been published in T he Journal of Clinical Investigation (Zhou, et al., 2006b), a high profile journal. It has long been known that inflammation and infection reduce expression of hepatic cytochrome P450 (CYP) genes involved in xenobiotic metabolism. Moreover, there is considerable evidence to suggest that exposure to xenobiotic chemicals can impair immune function. Yet the molecular mechanisms underlying both of these phenomena have remained largely unknown until our studies. We found that activation of SXR by commonly used drugs in humans leads to repression of target genes for the transcription factor NF-κB, a key regulator of inflammation and the immune response. Similarly, activation of SXR antagonizes NF-κB activity in cell culture and in vivo. NF-κB target genes are upregulated and small bowel inflammation is significantly increased in PXR knockout mice, suggesting that SXR antagonizes NF-κB signaling. Interestingly, NF-κB activation reciprocally inhibits SXR and expression of its target genes, whereas inhibition of NF-κB enhances SXR activity. This SXR–NF-κB axis provides a molecular explanation for the suppression of hepatic CYP mRNAs by inflammatory stimuli as well as the immunosuppressant effects of xenobiotics and SXR-responsive drugs. As such, it is extremely relevant to human health, both when considering xenobiotic metabolism and the potential risk of exposure to these compounds, and in molecular pharmacology.
An important unanswered question regarding the inflammation observed in PXR-/- mice (Zhou, et al., 2006b) is whether or not it is progressive, perhaps leading to cumulative tissue changes including cancer. This would be expected considering the increased levels of inflammatory mediators we observed in these animals (Zhou, et al., 2006b). Recently, we have undertaken more extensive tissue examination of six older (> 1 year old) PXR-/- mice. We noted that the livers and spleens are abnormal on gross examination, being more nodular in appearance and in the case of spleens having more prominent white pulp. This is reflected by the histologic evaluation of liver, spleen, and bowel (with accompanying pancreas in three of six animals). We selected representative photomicrographs of tissue sections from individual animals to demonstrate this observation (Figure 2). Although pathologic changes were observed in each of these tissues in essentially all animals, the range of disruption of normal tissue architecture is variable. Histologic examination of liver revealed varying amounts of mononuclear cell inflammatory infiltrates, identified by arrows, that could be rather diffuse and associated with extensive hepatocyte destruction and regeneration (Figure 2A) or more limited focal abnormal collections of mononuclear cells in the setting of slightly increased fatty change (Figure 2B). In several cases, these focal mononuclear cell aggregates were large enough to be visible with the naked eye, suggesting a true lymphoproliferative (cancerous) state. There was a greater degree of homogeneity in the spleens. A section from the central portion of one spleen (Figure 2C) and a longitudinal terminal section from another (Figure 2D) both show marked hypertrophy of the marginal zone, identified by the arrows, surrounding the periarteriolar sheath (PAS) region, identified by the arrowheads, of the white pulp. The former zone contains both T and B lymphocytes, whereas the PAS contains primarily T cells (CD8 > CD4), and red pulp has the largest density of macrophages. This histological pattern is consistent with the flow cytometry of wild type and PXR-/- splenocytes described below. Although pancreatic tissue was only identified in specimens obtained from three of the six animals, two of these demonstrated surprising pathology. Pancreatic and peri-pancreatic adipose tissues had focal mononuclear inflammatory infiltrates that, as with the liver specimens, could be associated with limited local pancreatic tissue destruction and cellular infiltrates (Figure 2E) or more extensive destruction and more diffuse and slightly less prominent mononuclear cell infiltration (Figure 2F). Finally, in addition to the observation of increased mucosal mononuclear inflammatory infiltrates, we observed a marked increase in Peyer’s patch size in five of six animals, with a histologic picture highly reminiscent of that seen in the spleen (Figure 2G, H). This suggests that it is likely that an identical cellular population is responsible. The observed tissue destruction in the liver and pancreas would be expected to result in recruitment of macrophages to sites of cell death. However, the observation of focal mononuclear infiltrates without associated cell death or tissue destruction argues that recruitment of macrophages by tissue destruction cannot wholly account for the increased mononuclear cell infiltrates observed in these diverse tissues of these six animals.

Figure 2. Inflammatory Lesions in Aged PXR-/- Mice. Hematoxylin and Eosin (H&E) stained representative sections of formalin fixed paraffin embedded tissues from 1.5 year old PXR-/- mice. Original magnification 40x. (A, B) Liver showing a range of pathology from widespread hepatocyte destruction, associated with mononuclear infiltration, to focal parenchymal mononuclear cellular inflammatory infiltrate. Inflammatory infiltrates are denoted by arrows. (C, D) Spleen sections from central and distal locations. Open arrowheads identify periarteriolar sheath regions and arrows identify the hypertrophic marginal zone. (E, F) Pancreas showing (E) a range of pathology extending from a focal region of destructive inflammation surrounded by preserved pancreas to (F) more widespread tissue destruction and inflammation with a faint rim of relatively preserved pancreatic exocrine tissue and ductile epithelium. (G, H) Cross-sections of proximal small bowel with hypertrophic Peyer’s patches.
As an initial and important step in understanding the molecular basis of this inflammation, we wish to determine which types of cells are predominantly found in the hypertrophic regions and whether these are monoclonal or polyclonal in origin. Initial observations revealed that the spleens of the PXR-/- mice were approximately 40% larger in size and mass than spleens from matched wild type mice. Single cell suspensions from wild type and PXR-/- spleens were examined by flow cytometry to begin to characterize the cellular components responsible for the abnormal histology noted in Figure 2. Interestingly, the most prominent finding is of a markedly increased B220 intermediate cell population in the PXR-/- mice relative to wild type. A differential analysis of markers associated with cells in the immune compartment leads us to conclude that the leading candidate for the increased cell populations in the chronically inflamed tissues (Figure 2) is the B1 B lymphocyte. These results are currently being written for publication and served as the basis for a recent National Institutes of Health (NIH) grant application, which will be required for future study of this phenomenon.
Organotins are Environmental Obesogens
Occasionally, novel and unexpected results lead to new research directions. During our characterization of the xenobiotic panel for its ability to activate SXR across species, we decided to also investigate whether these compounds activated other nuclear receptors. A surprising and striking result was that tributyltin (TBT) and triphenyltin were potent and efficacious activators of the vertebrate retinoid “X” receptors (RXR) and peroxisome proliferator activated receptor gamma (PPARγ) (Grun, et al., 2006). We and others recently showed that organotins act as high affinity agonistic ligands for RXRs and PPARγ. Exposure to environmentally relevant (nM) levels of TBT and related compounds activates RXRs and PPARγ, in cell culture and in vivo. TBT exposure in cell-based assays leads to differentiation of 3T3-L1 fibroblasts into adipocytes. A variety of molecular and cell-based studies confirms that TBT activates RXR and PPARγ to induce and regulate the expression of genes known to be involved in adipogenesis, in vivo. Mice exposed prenatally to TBT (but not thereafter) were more than 10 % heavier at 10 months of age, despite being reared on identical chow. Frogs exposed to nM levels of TBT or RXR activators showed replacement of gonadal tissue by adipocytes (Grun, et al., 2006). Taken together, our results establish the existence of environmental obesogens, chemicals that inappropriately regulate lipid metabolism and adipogenesis to promote obesity. The physiological consequences of receptor activation predict that RXR-heterodimer target genes and downstream signaling cascades are sensitive to organotin misregulation. Notably, the activation of RXR-PPARγ suggests that organotins should have profound effects on cellular differentiation programs, such as adipogenesis. In addition, effects on lipid/glucose metabolism, cholesterol homeostasis, steroid biosynthesis, and developmental morphogenesis may be expected (Grun, et al., 2006). These results are quite significant and have important public health implications. These studies are published and have led to the submission of several grant proposals.
Future Activities:
In the next year, we aim to complete the studies underway and publish the results. Specifically, we aim to accomplish the following:
- Publish our studies on species-specific responses of SXR to xenobiotic compounds, establishing the commonalities and differences in the response of the “SXR zoo” to ligands.
- Publish our studies on the effects of SNPs on the response of SXR to a panel of high-priority endocrine disrupting chemicals.
- Test the SNPs in human SXR for their ability to interact with the NF-κB signaling pathway and for the effects of the xenobiotic compounds above on this interaction, and publish the results.
- Publish our modeling studies describing a computational model for SXR agonists and antagonists.
- Publish our findings on the identification of SXR target genes.
- Publish our findings characterizing the “fully-humanized” SXR mouse.
- Publish our characterization of the inflammatory and immune phenotypes of the PXR-/- mouse.
Journal Articles on this Report : 8 Displayed | Download in RIS Format
| Other project views: | All 39 publications | 8 publications in selected types | All 8 journal articles |
|---|
| Type | Citation | ||
|---|---|---|---|
|
|
Grun F, Watanabe H, Zamanian Z, Maeda L, Arima K, Cubacha R, Gardiner DM, Kanno J, Iguchi T, Blumberg B. Endocrine-disrupting organotin compounds are potent inducers of adipogenesis in vertebrates. Molecular Endocrinology 2006;20(9):2141-2155. |
CR830686 (2006) |
Exit |
|
|
Grun F, Blumberg B. Environmental obesogens: organotins and endocrine disruption via nuclear receptor signaling. Endocrinology 2006;147(6):S50-S55. |
CR830686 (2006) |
Exit |
|
|
Gunderson MP, Kohno S, Blumberg B, Iguchi T, Guillette Jr LJ. Up-regulation of the alligator CYP3A77 gene by toxaphene and dexamethasone and its short term effect on plasma testosterone concentrations. Aquatic Toxicology 2006;78(3):272-283. |
CR830686 (2006) |
Exit Exit |
|
|
Tabb MM, Kholodovych V, Grün F, Zhou C, Welsh WJ, Blumberg B. Highly chlorinated PCBs inhibit the human xenobiotic response mediated by the steroid and xenobiotic receptor (SXR). Environmental Health Perspectives 2004;112(2):163-169. |
CR830686 (2003) CR830686 (2006) R826133 (Final) |
not available |
|
|
Tabb MM, Blumberg B. New modes of action for endocrine-disrupting chemicals. Molecular Endocrinology 2006;20(3):475-482. |
CR830686 (2006) |
Exit Exit |
|
|
Zhou C, Tabb MM, Sadatrafiei A, Grün F, Blumberg B. Tocotrienols activate the steroid and xenobiotic receptor, SXR, and selectively regulate expression of its target genes. Drug Metabolism and Disposition 2004;32(10):1075-1082. |
CR830686 (2004) CR830686 (2006) |
not available |
|
|
Zhou C, Tabb MM, Nelson EL, Grun F, Verma S, Sadatrafiei A, Lin M, Mallick S, Forman BM, Thummel KE, Blumberg B. Mutual repression between steroid and xenobiotic receptor and NF-κB signaling pathways links xenobiotic metabolism and inflammation. The Journal of Clinical Investigation 2006b;116(8):2280-2289. |
CR830686 (2006) |
Exit |
|
|
Zhou C, Assem M, Tay JC, Watkins PB, Blumberg B, Schuetz EG, Thummel KE. Steroid and xenobiotic receptor and vitamin D receptor crosstalk mediates CYP24 expression and drug-induced osteomalacia. The Journal of Clinical Investigation 2006a;116(6):1703-1712. |
CR830686 (2006) |
Exit Exit |
Supplemental Keywords:
xenobiotic, species-specific response, pollution, pesticide, inflammation, NF-κB,, Health, Scientific Discipline, ENVIRONMENTAL MANAGEMENT, Health Risk Assessment, Risk Assessments, Environmental Chemistry, Biochemistry, Risk Assessment, xenobiotics, human exposure, animal model, chemical exposure, steroids, human health risk, analysis of chemical exposureProgress and Final Reports:
Original AbstractThe perspectives, information and conclusions conveyed in research project abstracts, progress reports, final reports, journal abstracts and journal publications convey the viewpoints of the principal investigator and may not represent the views and policies of ORD and EPA. Conclusions drawn by the principal investigators have not been reviewed by the Agency.