
Grantee Research Project Results
2009 Progress Report: Endothelial Cell Responses to PM—In Vitro and In Vivo
EPA Grant Number: R832414C002Subproject: this is subproject number 002 , established and managed by the Center Director under grant R832414
(EPA does not fund or establish subprojects; EPA awards and manages the overall grant for this center).
Center: UC Davis Center for Children's Environmental Health and Disease Prevention
Center Director: Van de Water, Judith
Title: Endothelial Cell Responses to PM—In Vitro and In Vivo
Investigators: Wilson, Dennis , Rutledge, John
Current Investigators: Wilson, Dennis , Barakat, Abdul , Anastasio, Cort , Tablin, Fern , Rutledge, John
Institution: University of California - Davis
EPA Project Officer: Chung, Serena
Project Period: October 1, 2005 through September 30, 2010 (Extended to September 30, 2011)
Project Period Covered by this Report: July 1, 2008 through June 30,2009
RFA: Particulate Matter Research Centers (2004) RFA Text | Recipients Lists
Research Category: Human Health , Air
Objective:
Progress Summary:
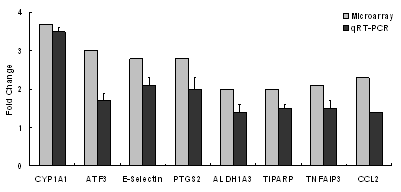
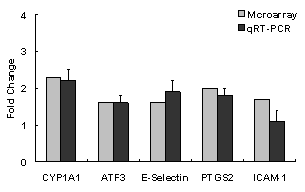
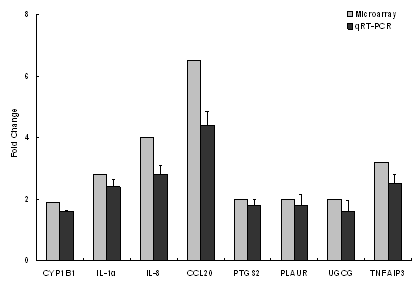


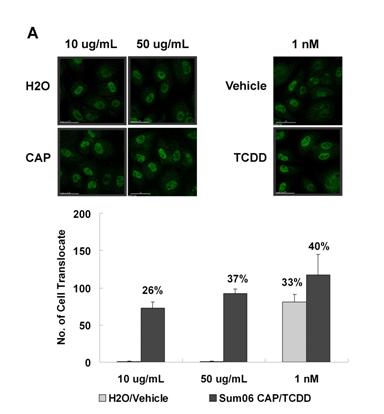

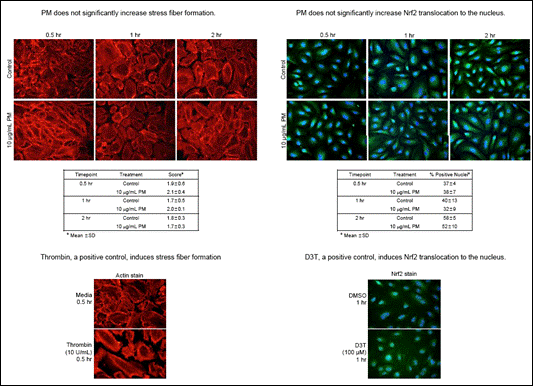
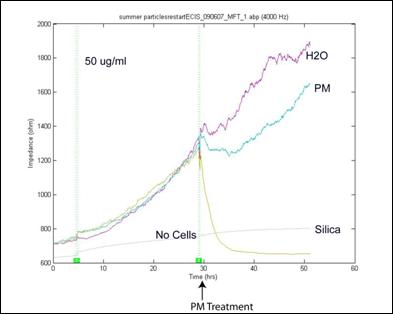
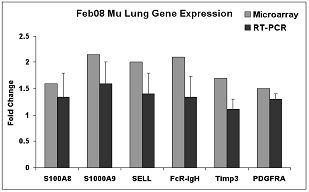
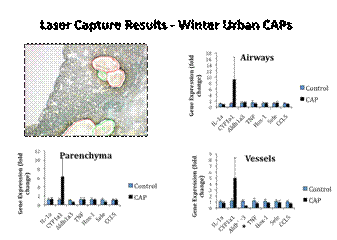
|
|
mRNA Expression (fold change from water control)
|
|||
|
|
PM (50 µg/mL)
|
SiO2 (50 µg/mL)
|
||
|
|
THP-1 monocytes
|
Fresh monocytes
|
THP-1 monocytes
|
Fresh monocytes
|
|
TNFα
|
2.67*
|
3.6*
|
4.3*
|
1.24
|
|
IL-1β
|
8.92*
|
4.82*
|
2.68*
|
1.57*
|
|
IL-6
|
34.92*
|
1.23
|
7.61*
|
1.97*
|
|
IL-8
|
8.73*
|
10.54*
|
2.46*
|
2.54*
|
|
COX-2
|
2.77*
|
3.53*
|
2.48
|
1.81*
|
|
GM-CSF
|
16.47*
|
5.79*
|
6.21
|
1.63
|
|
NFκB
|
1.89*
|
2.25*
|
0.98
|
0.94
|
|
IL-10
|
0.58
|
1.07
|
0.70
|
1.23
|
|
IL-12
|
0.43**
|
1.15
|
1.21
|
4.89*
|
|
CCL3 (MIP-1α)
|
0.61
|
1.71
|
1.58
|
1.14
|
|
CCL5 (Rantes)
|
0.99
|
1.12
|
2.15
|
1.61*
|
|
CYP1a1
|
3.21*
|
26.54*
|
|
3.24
|
|
Aldh3a1
|
0.74
|
1.08
|
|
2.52
|
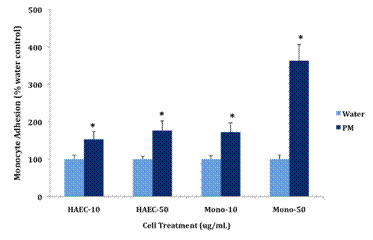


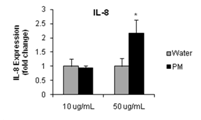
Future Activities:
Journal Articles:
No journal articles submitted with this report: View all 15 publications for this subprojectSupplemental Keywords:
Health, RFA, Scientific Discipline, Air, Health Risk Assessment, Risk Assessments, particulate matter, Environmental Chemistry, Epidemiology, ambient aerosol, lung disease, human exposure, long term exposure, airborne particulate matter, concentrated air particles, ambient particle health effects, epidemiological studies, PM, inhalation toxicology, toxicology, vascular dysfunction, acute cardiovascular effects, cardiovascular disease, human health risk, lung injury, microarray studies, airway diseaseProgress and Final Reports:
Original AbstractMain Center Abstract and Reports:
R832414 UC Davis Center for Children's Environmental Health and Disease Prevention Subprojects under this Center: (EPA does not fund or establish subprojects; EPA awards and manages the overall grant for this center).
R832414C001 Project 1 -- Pulmonary Metabolic Response
R832414C002 Endothelial Cell Responses to PM—In Vitro and In Vivo
R832414C003 Project 3 -- Inhalation Exposure Assessment of San Joaquin Valley Aerosol
R832414C004 Project 4 -- Transport and Fate Particles
R832414C005 Project 5 -- Architecture Development and Particle Deposition
The perspectives, information and conclusions conveyed in research project abstracts, progress reports, final reports, journal abstracts and journal publications convey the viewpoints of the principal investigator and may not represent the views and policies of ORD and EPA. Conclusions drawn by the principal investigators have not been reviewed by the Agency.