
Grantee Research Project Results
2016 Progress Report: A Bioenergetics-Based Approach to Understanding and Predicting Individual- to Community-Level Ecological Effects of Manufactured Chemicals
EPA Grant Number: R835800Title: A Bioenergetics-Based Approach to Understanding and Predicting Individual- to Community-Level Ecological Effects of Manufactured Chemicals
Investigators: J. Salice, Christopher
Institution: Towson University
EPA Project Officer: Aja, Hayley
Project Period: September 1, 2015 through August 31, 2018 (Extended to February 28, 2020)
Project Period Covered by this Report: September 1, 2015 through December 15,2016
Project Amount: $374,510
RFA: Systems-Based Research for Evaluating Ecological Impacts of Manufactured Chemicals (2014) RFA Text | Recipients Lists
Research Category: Chemical Safety for Sustainability
Objective:
A fundamental goal in ecotoxicology and ecological risk assessment (ERA) is to predict the frequency and magnitude of adverse ecological effects resulting from chemical contaminants. While there is a considerable volume of research generated on the effects of a wide variety of manufactured chemicals on ecologically relevant receptors, the vast majority of studies are focused at levels of biological organization that are most conducive to empirical approaches- the individual and lower. Paradoxically, the levels of biological organization that are most relevant to environmental health and societal value are at the population level and higher (communities and ecosystems). The overarching objective of the proposed research is to develop a bioenergetics-based Adverse Outcome Pathway framework that can translate from individual-level to population- and community-level effects of manufactured chemicals.
Progress Summary:
| Specific Aim 1: Establish bioenergetics-based responses and associated mathematical models (e.g., Dynamic Energy Budget (DEB), Von Bertelanffy growth model) of individual study species exposed to PFOS and pyraclostrobin for 2-4 week exposures, which are representative of relatively low-cost, standardized toxicity tests. Endpoints: Life history traits, cellular energy assimilation, metabolic rate, feeding rate, fatty acid profiles growth rate as a key endpoint Models: Single species bioenergetics (DEB) and growth models Summary Specific Aim 1 Progress: We have successfully conducted relatively short duration toxicity studies to explore the effects of pyraclostrobin and perfluorooctane sulfonate (PFOS) on Daphnia magna and/or Lymnaea stagnalis. Some results are provided below for PFOS. We are currently measuring lipid profiles as a bioenergetics endpoint with results forthcoming. We have also developed several models including DEB-IBM models for D. magna and L. stagnalis, cohort-based population models for D. magna as well as count-based population viability models for D. magna. Specific Aim 2: Establish bioenergetics-based responses, life-history effects and population-level effects of individual study species exposed to PFOS and pyraclostrobin using a Life Table Response Experiment study design (full life cycle exposure). Endpoints: Same as Specific Aim 1 plus population-level endpoints (population growth rate, estimated abundance, stable stage distribution) Models: Individual-based and cohort-based population models Summary Specific Aim 2 Progress: We have successfully conducted full life cycle studies involving D. magna exposed to both PFOS and pyraclostrobin and have developed and implemented a new study design in which D. magna populations are exposed to a toxicant and the population is followed through time. This design more closely represents how daphnia would be exposed in field conditions. The daphnia DEB-IBM does an excellent job of capturing D. magna population dynamics in this exposure scenario. We have also conducted toxicity studies with L. stagnalis but this species is not particularly sensitive to Pyraclostrobing or PFOS under our exposure conditions. Nonetheless, we have started working with multi-species systems that include D. magna and L. stagnalis and have observed some interesting dynamics. Specific Aim 3: Establish bioenergetics-based responses, life history effects and population-level effects of individual study species exposed to PFOS and pyraclostrobin in addition to the presence of conspecific competitors and predators. Endpoints: Same as Specific Aim 1 plus modified life history based on species interactions (competition coefficients; predator-prey coefficients) Models: Density dependent, resource based population models; Modified population models that include mortality from predation Summary Specific Aim 3 Progress: We have initiated studies in which we have combined L. stagnalis and D. magna. In this scenario, the nature of the interaction is energetic facilitation to D. magna via L. stagnalis. Specifically, L. stagnalis "liberates" sources of energy (lettuce) making it available to D. magna. This is a relatively unstudied form of species interaction but one that is likely common and potentially important. Also, our population study design for D. magna includes density-dependent effects and hence, intra-species competition. Our DEB-IBM also captures these dynamic well and illustrates one important attribute of IBMs - properties and dynamics of populations can emerge from the interactions and behaviors of individuals. EXAMPLE RESULTS AND UPDATES: Ecotoxicity of PFOS to L. stagnalis and D. magna PFOS is not toxic to Lymnaea stagnalis with toxicity occurring far above environmentally relevant concentrations (> 3 mg/L (ppm)). I report only effects on survival of adults. Adult snails were exposed to 0, 1.5, 3, 6, 12.5, 25, and 50 ppm and effects on survival were observed in the 6 (p = 0.03), 25 (p < 0.001), and 50 (p = 0.03) µg/mL concentrations which had significantly decreased survival when compared to the controls. These effect levels are several orders of magnitude greater than typical, high-end environmental concentrations. Figure 1. Kaplain-meier curves for survival of adult snails exposed to PFOS. Each concentration is represented by three colored lines. The middle line represents the number of snails alive on each day of the study, and the upper and lower lines are the 95% confidence boundaries. Similar to results of PFOS toxicity studies with L. stagnalis, D. magna are not very sensitive to PFOS. However, there was concern that typical experimental exposure durations (21 days or less) are not representative of potential temporal exposure profiles for PFOS which appear to be at very low concentrations (single digit ppb) but over an extended period of time. Hence, we conducted experiments to explore this exposure profile in D. magna. Using a novel experimental design in which laboratory populations of D. magna were followed for several generations, we examined responses to relevant concentrations of PFOS which were 5 and 10 ppb. Figure 2 show the population counts through time for control, 5 ppb and 10 ppb PFOS-exposed Daphnia populations. All three treatments show similar population dynamics. However, to better analyze these data, we used count-based population viability analyses and estimated time to quasi-extinction (Figure 3). Interestingly, these analyses point to negative effects of PFOS on extinction risk.
Figure 2. Population-level responses of D. magna to PFOS at environmentally relevant exposure concentrations and durations. There were no strong, significant negative effects but there was greater variability in PFOS-exposed D. magna populations. Figure 3. Time to quasi-extinction in D. magna exposed to PFOS at environmentally relevant concentrations and durations. Results indicate greater extinction risk in populations exposed to PFOS. Bioenergetics-Based ModelingAn important objective of the project is to develop, test and vet different models to relate individual-level effects to effects at higher levels of biological organization. Because of significant advances in the use of Dynamic Energy Budget Models (DEB), we chose to develop DEB-Individual Based Models for D. magna and L. stagnalis. These models, in theory, can account for a wide range of effects and translate well to potential population level effects. We were successful in building Dynamic Energy Budget-Individual Based Models for both Daphnia magna and Lymnaea stagnalis. One student, Andrew East, M.S. (awarded December 2016) completed a thesis titled: A modeling framework to explore bioenergetic effects of environmental stress and interspecific interactions in aquatic invertebrates. His objective was to develop the D. magna and L. stagnalis DEB-IBMs and to further explore how the models could be used to understand and predict species interactions. Figure 4 provides an example of the output from the D. magna DEB-IBM compared to laboratory obtained population count data. Note the strong agreement between model results and data suggesting that this implementation of the DEB-IBM for D. magna has some strong predictive capacity under very controlled conditions.
Figure 4a, 4b, 4c, and 4d. Plots of experimental observations (top L and R) and model output (bottom L and R). Top left (a.) shows each replicate (connected points) of one liter populations through 40 days observed by the author. Top right (b.) shows additional datasets published by Martin, et al. (2013b) and Preuss, et al. (2009). Bottom left (c.) shows the current model plotted against all datasets. Bottom right (d.) shows an additional model output using different Neighborhood Effect model parameters. Solid lines are mean of 50 simulations and gray shading represents maximum and minimum. We are currently conducting additional experiments with pyraclostrobin but explored a theoretical implementation of the D. magna DEB-IBM that accounts for effects of a toxicant. In this case, adjusting the metabolic cost of detoxification causes a decrease in overall population counts and a slight shift in the timing of the population peak (Figure 5). Figure 5. Plot of three model population simulations under a range of stressor exposures. Solid lines are mean and dashed are maximum and minimum of 50 simulations. Stressor exposure was during the first three days and occurred every third day with a 2 day half-life. Exposure in each patch (6% or 12%) was random but normally distributed with a standard deviation of 0.5%. We also developed a DEB-IBM for L. stagnalis. Our research, however, has shown that L. stagnalis is generally much less sensitive to PFOS and Pyraclostrobin compared to daphnia and that its longevity (> 1 year) suggests that this species may not play a significant role with regard to toxicant effects. Alternatively, L. stagnalis does represent a significant community species by facilitating the movement of energy from unavailable sources (macrophytes; lettuce in experiments) to available (nutrients, bacteria) for D. magna. This is an interesting, understudied but potentially important inter-species interaction. As a result, we mostly focused on L. stagnalis growth, which was successfully modeled using a modified DEB model (Figure 6). We used a spatially-explicit, behavior-based IBM as well with spatial dimensions that reflect experimental containers (e.g., Figure 7 is a representation of the modeled environment). Figure 6. Juvenile assimilation lag model (red line) fit against growth data (black crosses) from Zimmer, et al. (2014). Final size data and parameters (Table 1) were used to estimate an f value of 0.85 and in combination with food quality parameter (a) and length at regular assimilation (Table 1) the model fits data well.
 Figure 7. Two simulated environment shapes used to simulate growth of snails. (a.) is shallow with an area of 796 cm3 and dimensions of 11x11x6 cm and (b.) is narrow with an area of 796 cm3 and dimensions of 11x6x11 cm. Green patched are single cm2 patches containing lettuce. We experimentally explored the interactions between D. magna and L. stagnalis via experimentation and coupled modeling. The Interactions Between Daphnia magna and Lymnaea stagnalis (Figure 8).
First, we confirmed that energetic facilitation of D. magna by L. stagnalis occurs and has significant impacts on D. magna life history and population dynamics. This is evident from the data provided in Figure 9 showing how the presence of snails causes D. magna populations to increase rapidly and reach the highest peak (Figure 9, ASLD). Figure 10 shows modeled D. magna growth and actual size data for the different facilitation treatments. Two important points are illustrated by Figure 10. First, the model does a good job of capturing D. magna growth under a range of different energetic environments. Second, there are clear and strong differences in growth dependent on the energetic environment. In particular, the SLD daphnia (exposed to snails and lettuce only (no algae) show growth trajectories that are markedly different than other groups. Figure 9. D. magna population size of each replicate and treatment through time. Replicates are points connected by lines. Each plot has the 'LD' treatment for consistent comparison. All data from day 40 has 10 individuals added to account for sampling of individuals on day 39.
Figure 10. Maximum size measured in each replicate prior to day 28. Fitted von Bertalanffy functions are plotted against data and growth rate parameters (k) are highlighted for each treatment. The model's performance across the three highlighted treatments (ASD, SLD, and ASLD) demonstrates its performance overall (Figure 11, 12, 13). Clearly, mean model output captured timing and size of daphnid populations quite well under a range of facilitation levels (ASD and ASLD). Additionally, the relatively simple quantitative methodology (snail waste converted to algae cells) appeared to explain the observed persistence of daphnid populations in the experimental treatment without amended algae (SLD).
Figure 11. Model output (red and black lines) against experimental output (blue points) for the ASD scenario. Dark lines are means and light lines are maximum and minimum. Model data are of 50 simulations with algae levels set to normal and lettuce to minimum.
We have conducted several toxicity studies involving multiple species (L. stagnalis and D. magna) and are assembling a database of lipid profiles for stressed species. We will be running more multi-species toxicity studies this year to include mesocosms with more than 2 species. We are measuring different lipid classes is organisms and will use this as an indicator of energetic disruption in toxicant-exposed organisms |
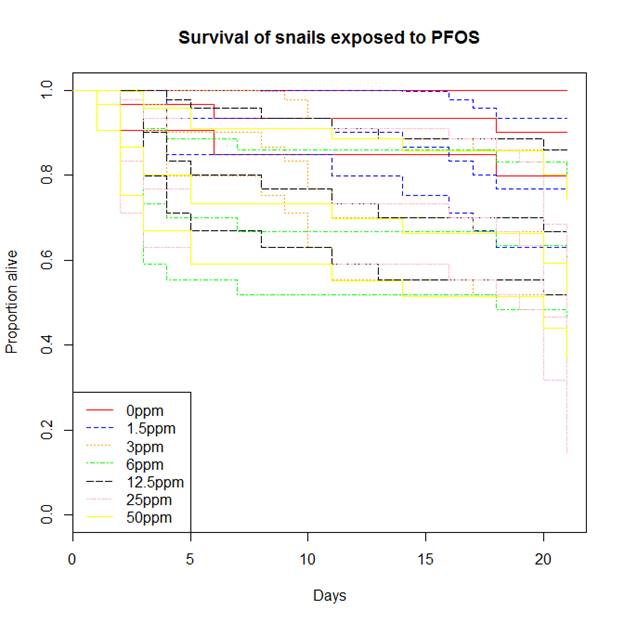
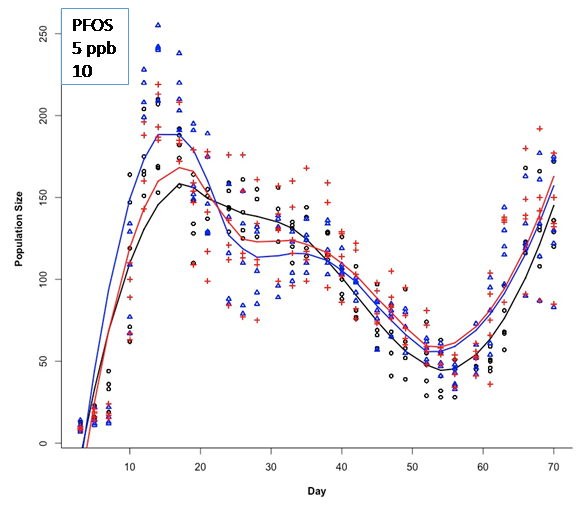
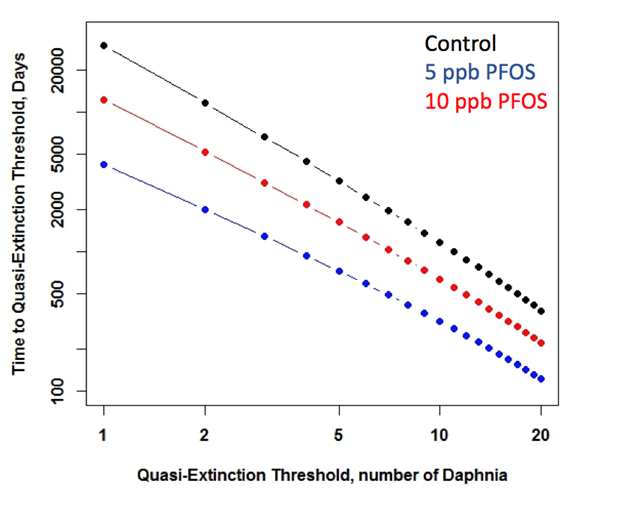
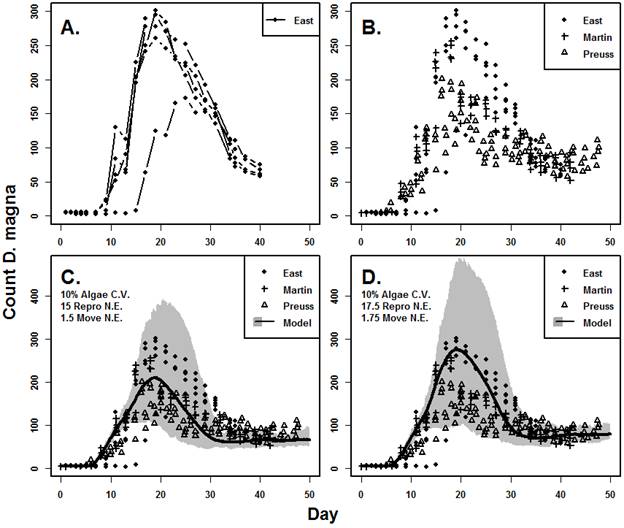
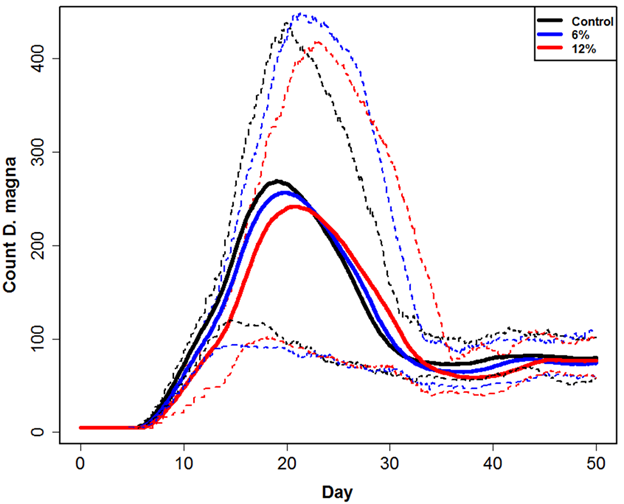
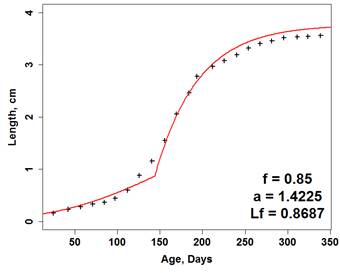
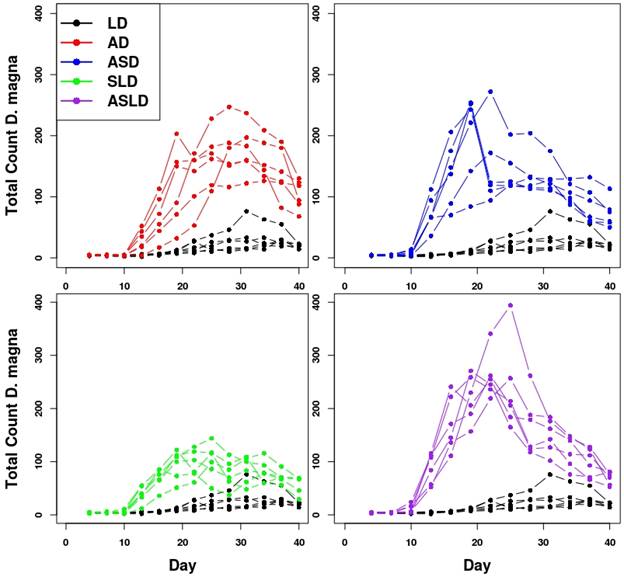
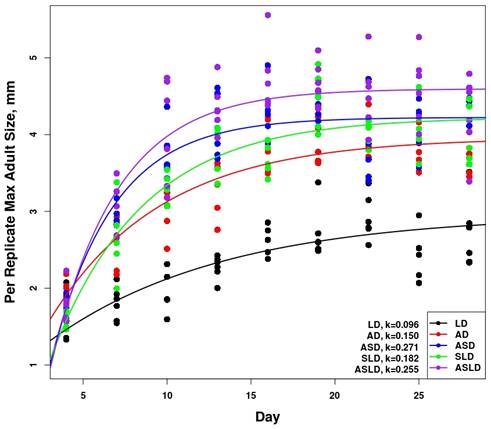
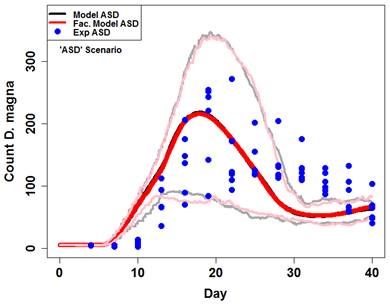
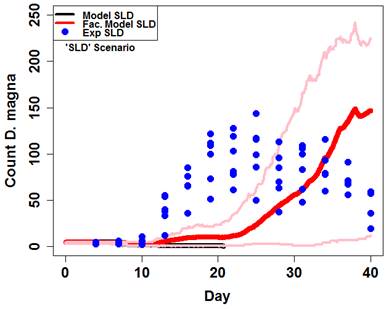 Figure 12. Model output (red and black lines) against experimental output (blue points) for the SLD scenario. Dark lines are means and light lines are maximum and minimum. Model data are of 50 simulations with algae levels set to normal and lettuce to minimum.
Figure 12. Model output (red and black lines) against experimental output (blue points) for the SLD scenario. Dark lines are means and light lines are maximum and minimum. Model data are of 50 simulations with algae levels set to normal and lettuce to minimum.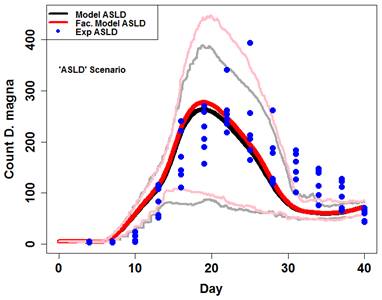 Figure 13. Model output (red and black lines) against experimental output (blue points) for ASLD scenario. Dark lines are means and light lines are maximum and minimum. Model data are of 50 simulations with algae levels set to normal and lettuce to minimum.
Figure 13. Model output (red and black lines) against experimental output (blue points) for ASLD scenario. Dark lines are means and light lines are maximum and minimum. Model data are of 50 simulations with algae levels set to normal and lettuce to minimum.Future Activities:
We will continue to develop and use a variety of mathematical models to understand and predict effects of toxicants on ecological receptors and systems. The DEB-IBM construct appears especially promising in being able to capture growth and population dynamics of study species. However, cohort-based and count-based population models also have been useful in understanding effects of multiple stressors and subtle effects on population counts. Our evolving view is that it is likely a multi-model perspective is useful and that the model construct is largely dependent on the data. We will continue to present at local, national and international meetings and have several manuscripts in preparation with related papers already published.
Journal Articles on this Report : 2 Displayed | Download in RIS Format
| Other project views: | All 26 publications | 9 publications in selected types | All 9 journal articles |
|---|
| Type | Citation | ||
|---|---|---|---|
|
|
Reategui-Zirena EG, Fidder BN, Olson AD, Dawson DE, Bilbo TR, Salice CJ. Transgenerational endpoints provide increased sensitivity and insight into multigenerational responses of Lymnaea stagnalis exposed to cadmium. Environmental Pollution 2017;224:572-580. |
R835800 (2016) R835800 (2017) R835800 (2018) R835800 (2019) |
Exit Exit Exit |
|
|
Rohr JR, Salice CJ, Nisbet RM. The pros and cons of ecological risk assessment based on data from different levels of biological organization. Critical Reviews in Toxicology 2016;46(9):756-784. |
R835800 (2016) R835800 (2017) R835800 (2018) R835800 (2019) R835188 (Final) R835797 (2015) R835797 (2016) |
Exit Exit |
Progress and Final Reports:
Original AbstractThe perspectives, information and conclusions conveyed in research project abstracts, progress reports, final reports, journal abstracts and journal publications convey the viewpoints of the principal investigator and may not represent the views and policies of ORD and EPA. Conclusions drawn by the principal investigators have not been reviewed by the Agency.