
Grantee Research Project Results
Final Report: ECOHAB: Trophic effects of two dinoflagellates.
EPA Grant Number: R826219Title: ECOHAB: Trophic effects of two dinoflagellates.
Investigators: McManus, George , Dam, Hans G. , Shumway, Sandra E. , Smolowitz, Roxanna M. , Wikfors, Gary H.
Institution: University of Connecticut , Long Island University - Southampton College , Marine Biological Laboratory , National Oceanic and Atmospheric Administration
EPA Project Officer: Packard, Benjamin H
Project Period: January 15, 1998 through January 14, 1999
Project Amount: $102,514
RFA: Harmful Algal Blooms (1997) RFA Text | Recipients Lists
Research Category: Aquatic Ecosystems , Water
Objective:
The objective of the project was to document, under controlled laboratory conditions, effects of two planktonic harmful algal bloom (HAB) dinoflagellates, Prorocentrum minimum (Milford Lab strain EXUV) and Gyrodinium aureolum, upon a taxonomically-diverse list of benthic and pelagic consumers: larval and post-set bay scallops, copepods, and micro-zooplankton (mainly ciliates). Our plan was to evaluate both feeding response and histological condition, especially of digestive organs, of animals fed unialgal and mixed algal diets that included the two dinoflagellates. We followed this plan closely, with the exception that we substituted the raphidophyte alga Heterosigma carterae (Milford Lab strain OL) for Gyrodinium aureolum. The latter grew much too slowly for the large number of experiments required for the project. Although we focused mainly on P. minimum, Heterosigma provided a good substitute for the second dinoflagellate because it grew well for us, showed harmful effects on some of the pelagic grazers that were not harmed by Prorocentrum, and has been an HAB species of increasing concern over the past several years. In addition to P. minimum and H. carterae, we performed some experiments with Gymnodinium splendens being fed upon by juvenile scallops. In continuing work not funded by EPA, we are looking for toxic effects of a different P. minimum strain, one that was isolated from a bloom in the York River, VA, and which appears to be more toxic than the strain we have been using.
Summary/Accomplishments (Outputs/Outcomes):
Pelagic consumers
Copepods. We tested the hypotheses that 1) P. minimum is toxic to copepods; 2) P. minimum is nutritionally deficient for copepods. We performed experiments in which adult females of the copepod A. tonsa isolated from Long Island Sound were fed: a) cells in exponential growth phase, b) cells in early exponential, late exponential and senescent growth phases (Fig. l), or c) mixtures of P. minimum and the diatom Thalassiosira weissflogii. The clone used (EXUV) was also isolated from Long Island Sound and has been shown to have toxic effects on larvae and juvenile stages of oysters and clams from Long Island Sound (Wikfors and Smolowitz, 1993, 1995). In each case we measured ingestion, egg production and egg hatching rates at food (carbon) concentrations ranging from limiting to saturating conditions (experiments a and b); or at a limiting fixed carbon concentration (approx. 250 µgC L-l), but with different proportions (0,25,50,75,100%) of in the mixed diet (experiment c).
Experiments a and b were designed to obtain information on the functional and numerical responses of A. tonsa and whether these responses varied with the growth stage of the cells. These experiments revealed that A. tonsa readily ingests cells-up to the equivalent of > 200% of body carbon/day (Fig. 2), but that egg production is relatively low -maximum egg production rate < 20% of body carbon/day (Fig. 3). Hence, the gross growth efficiency was low and averaged 6% (Fig. 4). There was no indication that senescence of the cells resulted in lower ingestion or egg production rates relative to cells actively growing (Fig. 5,6). Egg hatching rates were high (> 8O%j and independent of the cell growth phase (Fig. 7). The mixture experiments (c) were designed to test the toxicity hypothesis following the recommendations of Jonasdottir et al. (1998) and revealed no evidence of toxicity for (Fig. 8). We conclude that represents a nutritionally deficient (as evidenced by the low growth efficiency), but not a toxic, diet for A. tonsa. Our results, which are in strong contrast to those of Wikfors and Smolowitz (1993, 1995), suggest that the impact of HABs throughout the food-web is strongly dependent on the composition of the consumers.
Ciliates. We examined the effects of P. minimum and H. carterae on growth rates of several species of choreotrich ciliates. This subclass comprises the largest group of pelagic herbivores among the Protista. Quantitatively, they are probably the most important primary herbivores in the sea (Pierce and Turner 1992). Because they are difficult to cultivate over more than a few hundred generations, we isolated them from Long Island Sound and grew them as needed for individual experiments (Gifford 1985).
For a typical experiment with P. minimum, ciliates were picked from cultures grown on mixed phytoplankton as food, and acclimated for 1 day at different concentrations of P. minimum in 10 ml wells. We then transferred ciliates to fresh wells at various concentrations of P. minimum, in triplicate at each concentration, and measured growth rates subsequently over 4-6 days. Results of three such experiments are shown in figure 9. Five out of six isolates tested showed good growth on P. minimum. One reason we tested over a wide range of algal concentrations is because we were interested in two different questions: could ciliates use this species effectively as food at low concentrations (i.e. under pre-bloom conditions in the field), and is there any inhibitory effect of the alga at very high concentrations (i.e. during a bloom)? For the ciliates that were able to grow on P. minimum as a sole food source, growth vs. food concentration curves showed typical behavior, saturating at 103 104 cells/ml. Above this level, the ciliates were growing at their maximum rates, and showed no evidence of inhibition, even at levels only rarely if ever encountered in nature. In fact, P. minimum is apparently a good food for some choreotrichs, many of which require mixtures of several species as food before they will grow. Results with the tintinnid Tintinnopsis tubulosoides (Fig. 9, lowest panel) are interesting because they appear to show not only an inability to grow-on this species, but higher mortality under bloom-like conditions.
To date, we have examined effects of Heterosigma carterae on only two species of ciliate. In both cases, ciliates did not grow when fed this alga as a sole food. In addition, qualitative observations indicated some possible sublethal effects, including cessation of swimming in the presence of Heterosigma.
Because it would be difficult or impossible to cultivate and perform feeding experiments on all of the ciliates commonly found in local waters, we cannot generalize from these few isolates about the overall role of ciliates in grazing down existing blooms or preventing them from occurring through their feeding activities. We thus used our EPA support to explore the possibility of studying grazing on HAB species by intact natural communities of ciliates. The best approach to this is to use some kind of marker to label the HAB species of interest (e.g. P. minimum) and introduce it to field-collected samples containing a ciliate assemblage (cf. (Rublee and Gallegos 1989)). Although our results are preliminary, we have been successful in using the DNA fluorochrome SYBR Green I to stain live P. minimum cultures and feed them to ciliates. If the technique proves robust, we could use it to evaluate the role of ciliates in grazing HABs at various stages of bloom development in the field. Preliminary results of these observations have been presented at one regional meeting, and further experiments on this technique are ongoing (McManus 1999).
Benthic consumers
We focused on the benthic consumer Argopecten irradians, the bay scallop. Our experiments involved feeding trials with larvae and post-set larvae, as well as juveniles. We also exposed embryos to both P. minimum and H, carterae to evaluate whether toxicity of either HAB species could be expressed through soluble toxins, In addition to measuring mortality, we evaluated whether feeding stages could ingest these species, whether they deposited filtered cells in "pseudofeces" rather than ingesting them, and whether sublethal behaviors initiated by the algae could be observed. Our results, including some observations on two other HAB isolates, a Gymnodinium splendens and a P. minimum isolate from the York River, VA, are summarized in table 1. Samples for post-set larvae and juveniles have been prepared for histological analysis and examination of these is ongoing.
As Table 1 indicates, some planned experiments with embryos and larvae were not conducted; this was because availability of larvae at the Milford Lab was lower than expected. By contrast, experiments with scallop juveniles exposed to HAB algae were expanded to include two additional algal strains - Gymnodinium splendens and a new isolate of Prorocentrum minimum from the York River (Chesapeake Bay) -- and to include flow-cytometer studies of feeding behavior with mixtures of HAB algae and control algal cultures (e.g. the prasinophyte Tetraselmis sp., a standard food in shellfish aquaculture). Only the York River P. minimum was acutely toxic to juvenile scallops, but mortality was rapid (18 hr) and thorough (100%). Flow-cytometer examination of juvenile scallop feeding behavior in mixtures of HAB and control algae showed that some HAB algae (P. minimum and H. carterae) were filtered "preferentially" and rejected as pseudofeces, after which the control alga was filtered and ingested. One very large dinoflagellate, Gymnodinium splendens, was only filtered after control-alga densities had been grazed down. Overall findings suggest that scallops actively remove HAB algae from suspension, but reject these cells (usually still living) in loose, pseudofeces strands. Thus, at least for short-term exposures, scallops serve to transfer HAB algae from pelagic to benthic compartments. Follow-up studies will include longer-term exposures to nearly-continuous mixtures of HAB and control algae to see if there is a point at which the scallops are no longer able to "protect" themselves by selective rejection of HAB algae.
| Alga | Embryos | Larvae | Post-larvae | Juveniles |
| P. minimum | No effect | Prevent setting | 3-day exposure histology | Mortality (days), pseudofeces (1998)1999 -- no mortality 105 cells/ml for 5 d filtered 1st from mix, rejected in pseudofeces. Histology |
| G. aureolum | 105cells/ml for 18 hr: no mortality histology | |||
| H. carterae | No effect | No apparent effect | Pseudofeces, valve "clapping" 104 cells/ml for 3 d; no mortality; H. carterae filtered 1st from mix, rejected in pseudofeces. Histology | |
| Gymnodinium splendens | 103 cells/ml for 5 d; no mortality Tetraselmis (control food) filtered 1st from mix, then Gymnodinium splendens. Histology | |||
| P. minimum,York River | 105 cells/ml for 18 hr; 100% mortality. |
| Figure 1. Percent sands + fines in wadeable streams of the Kilchis and Tillamook sub-basins | Figure 2. Large wood (m3/m2) in streams of the Kilchis and Tillamook sub-basins |
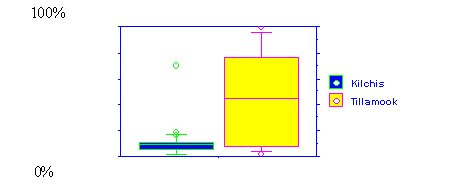 | 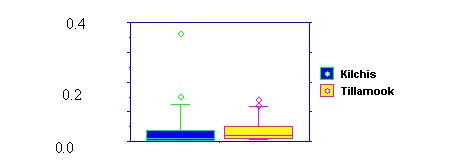 |
| Figure 3. Percent salmonids in summer low-flow fish assemblages of streams in the Kilchis and Tillamook sub-basins | FIgure 4. Acartia tonsa: Egg Production Versus Ingestion |
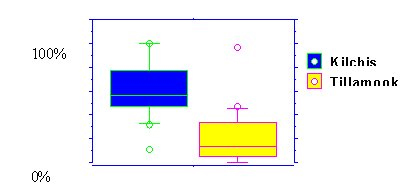 | |
| Figure 5. A. tonsa: Ingestion of P. minimum at different stages of the bloom | Figure 6. A. tonsa: Egg production during different stages of bloom |
| Figure 7. Acartia tonsa: hatching success at different stages of P. minimum bloom | Figure 8. Acartia tonsa Egg production vs. percent Thalassiosira weissfloggii in diet |
Figure 9.
Conclusions:
Our results are consistent with the idea that blooms of P. minimum have different effects on benthic vs. pelagic grazers. Our experiments with copepods and ciliates (pelagic consumers) did not show strong harmful effects of this HAB species. The benthic grazer, on the other hand, did show both lethal and sublethal effects of exposure to P. minimum, in addition to behavioral responses that suggested avoidance of ingestion. These results suggest a scenario in which blooms of P. minimum may originate in shallow bays where they can outgrow the grazing pressure of benthic consumers such as Argopecten, with subsequent export of bloom-level concentrations of the alga to the open water, where pelagic grazers dominate. Further studies, including especially fieldwork on naturally occurring blooms or mesocosm experiments with mixtures of benthic and pelagic consumers, are needed to test this hypothesis. Observations on Heterosigma carterae confirm results of earlier investigators who found this species to be unpalatable to consumers (e.g. Egloff 1986).
Journal Articles on this Report : 4 Displayed | Download in RIS Format
| Other project views: | All 9 publications | 4 publications in selected types | All 4 journal articles |
|---|
| Type | Citation | ||
|---|---|---|---|
|
|
May S, Burkholder J, Shumway S, Hegaret H, Wikfors G, Frank D. Effects of the toxic dinoflagellate Alexandrium monilatum on survival, grazing and behavioral response of three ecologically important bivalve molluscs. HARMFUL ALGAE 2010;9(3):281-293. |
R826219 (Final) R831704 (Final) |
Exit Exit |
|
|
Colin SP, Dam HG. Latitudinal differentiation in the effects of the toxic dinoflagellate Alexandrium spp. on the feeding and reproduction of populations of the copepod Acartia hudsonica. HARMFUL ALGAE 2002;1(1):113-125. |
R826219 (Final) |
Exit |
|
|
Dam HG, Colin SP. Prorocentrum minimum(clone Exuv) is nutritionally insufficient, but not toxic to the copepod Acartia tonsa. HARMFUL ALGAE 2005;4(3):575-584. |
R826219 (Final) |
not available |
|
|
Colin SP, Dam HG. Testing for resistance of pelagic marine copepods to a toxic dinoflagellate. EVOLUTIONARY ECOLOGY 2005;18:355-377. |
R826219 (Final) |
not available |
Supplemental Keywords:
estuary, fish kill, saxitoxin, oceanography, coastal, HPLC., RFA, Scientific Discipline, Ecosystem Protection/Environmental Exposure & Risk, Water, algal blooms, Ecological Indicators, Ecosystem Protection, Biology, Ecosystem/Assessment/Indicators, Oceanography, exploratory research environmental biology, Ecological Effects - Environmental Exposure & Risk, Ecology and Ecosystems, ECOHAB, phytoplankton, growth cycle, protozoans, coastal ecosystem, trophic transfer of phycotoxins, trophic effects, benthic algae, bloom dynamics, ecological exposure, dinoflagellate, dinoflagellates, fish kills, harmful algal blooms, copecod grazingThe perspectives, information and conclusions conveyed in research project abstracts, progress reports, final reports, journal abstracts and journal publications convey the viewpoints of the principal investigator and may not represent the views and policies of ORD and EPA. Conclusions drawn by the principal investigators have not been reviewed by the Agency.