
Grantee Research Project Results
2019 Progress Report: A Bioenergetics-Based Approach to Understanding and Predicting Individual- to Community-Level Ecological Effects of Manufactured Chemicals
EPA Grant Number: R835800Title: A Bioenergetics-Based Approach to Understanding and Predicting Individual- to Community-Level Ecological Effects of Manufactured Chemicals
Investigators: J. Salice, Christopher
Institution: Towson University
EPA Project Officer: Aja, Hayley
Project Period: September 1, 2015 through August 31, 2018 (Extended to February 28, 2020)
Project Period Covered by this Report: January 1, 2019 through December 15,2019
Project Amount: $374,510
RFA: Systems-Based Research for Evaluating Ecological Impacts of Manufactured Chemicals (2014) RFA Text | Recipients Lists
Research Category: Chemical Safety for Sustainability
Objective:
A fundamental goal in ecotoxicology and ecological risk assessment (ERA) is to predict the frequency and magnitude of adverse ecological effects resulting from chemical contaminants. While there is a considerable volume of research generated on the effects of a wide variety of manufactured chemicals on ecologically relevant receptors, the vast majority of studies are focused at levels of biological organization that are most conducive to empirical approaches – the individual and lower. Paradoxically, the levels of biological organization that are most relevant to environmental health and societal value are at the population level and higher (communities and ecosystems). The overarching objective of the proposed research is to develop a bioenergetics-based Adverse Outcome Pathway framework that can translate from individual level to population and community level effects of manufactured chemicals
Progress Summary:
Over the last year we have continued exploring effects of complex ecological conditions on Daphnia magna laboratory population dynamics. We have experimentally determined that specifics of the resource environment have significant impacts on the response of daphnids to environmental toxicants. Specifically, is appears that energetically available carbon has a stronger effect on toxicant resistance than does unavailable organic carbon that may sequester organic toxicants. Ongoing research exploring the impacts of nonconsumptive predator effects show a complex array of effects dependent on specifics of the exposure environment as well as the presence of other stressors. Here again, carbon that is contributed by predators (waste) may have consequences for prey. This mirrors previous work where we showed that waste generated by freshwater snails can have positive effects on Daphnia magna through what we called “energetic facilitation." The final experiments are being completed and will be presented and summarized in the final report. Overall, the research project has been very successful and has honed our understanding of how organismal and environmental energetics influence organismal responses to environmental stress.
Progress Summaries by Aims:
Specific Aim 1: Establish bioenergetics-based responses and associated mathematical models (e.g., Dynamic Energy Budget (DEB), Von Bertelanffy growth model) of individual study species exposed to PFOS and pyraclostrobin for 2-4 week exposures, which are representative of relatively low-cost, standardized toxicity tests.
Endpoints: Life history traits, cellular energy assimilation, metabolic rate, feeding rate, fatty acid profiles – growth rate as a key endpoint
Models: Single species bioenergetics (DEB) and growth models
Summary Specific Aim 1 Progress: We have continued exploring the effects of the resource environment and multiple stressors primarily on Daphnia magna. Undergraduate student, Maddie Barry, has conducted a series of experiments exploring whether the effects of the resource environment on decreasing toxicity are mediated through bioenergetic processes or via physico-chemical processes.
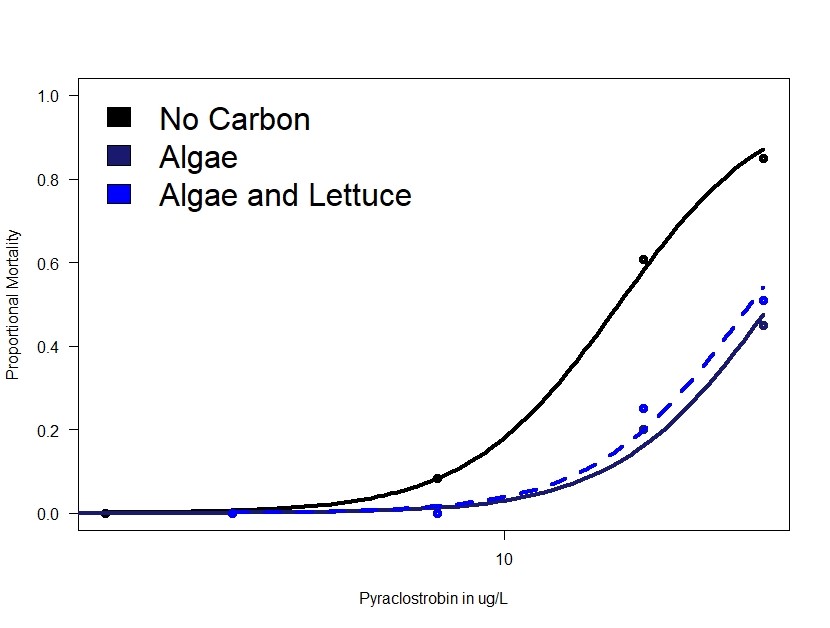
Figure 1 shows the influence of carbon presence and type on response to pyraclostrobin toxicity in Daphnia magna. The key result is that a lack of carbon increases toxicity of pyraclostrobin. We have now observed this many times and hypothesize that this mechanism occurs because of an energetic benefit of the carbon (algae) or as a result of sequestration of pyraclostrobin (any organic carbon) that reduces exposure.
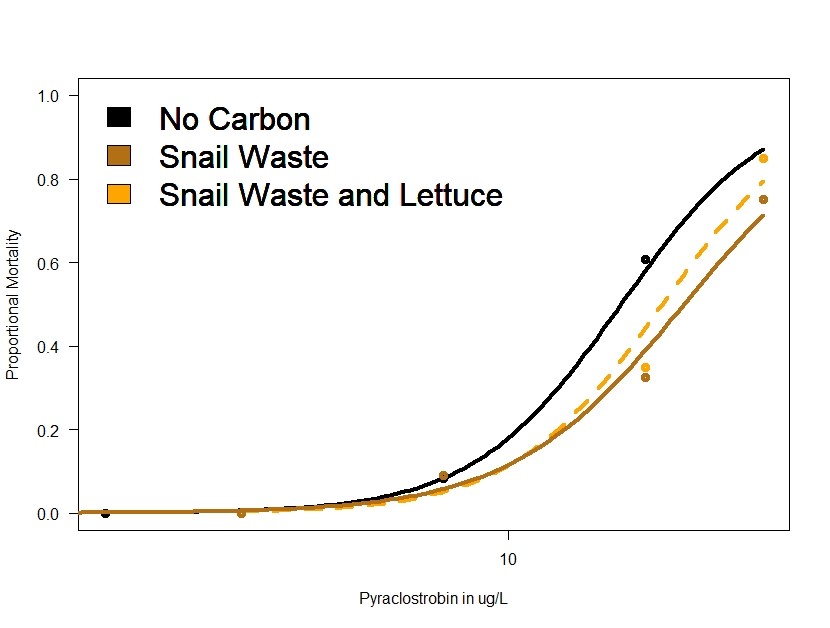
Figure 2 provides further evidence that the toxicity of pyraclostrobin is reduced in the presence of carbon, although in this case, the carbon sources are not energetically useful to daphnia and the effect of these carbon forms is less than algae (an energetically available carbon source).
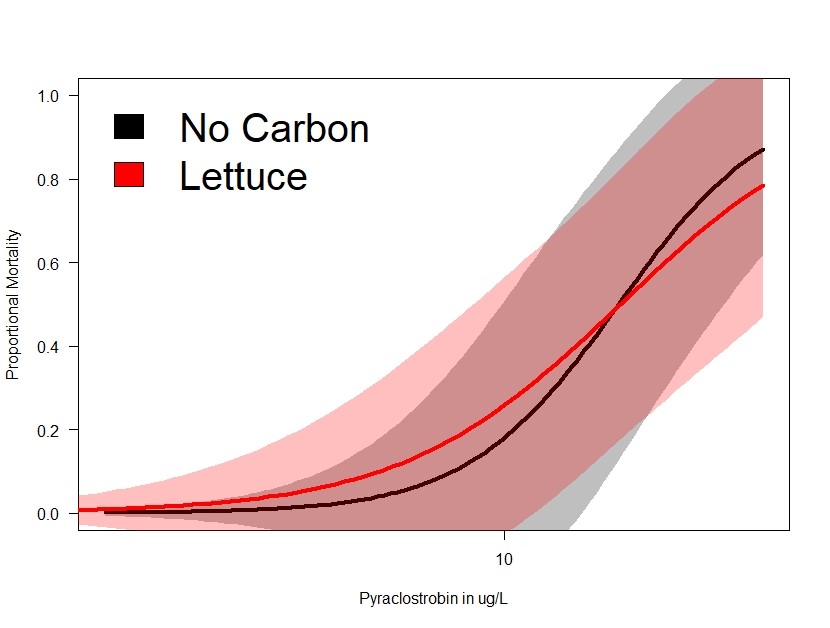
Figure 3 essentially shows that the primary beneficial effect of carbon source on reducing toxicity of pyraclostrobin is likely an energetic effects. Lettuce is an organic carbon source that can interact with organic contaminants like pyraclostrobin but is not a suitable energy source for daphnia in the short term. Because there is no difference in toxicity between lettuce-only and no carbon, this suggests that the reduced toxicity in algae fed daphnia is the result of an energetic advantage to daphnia.
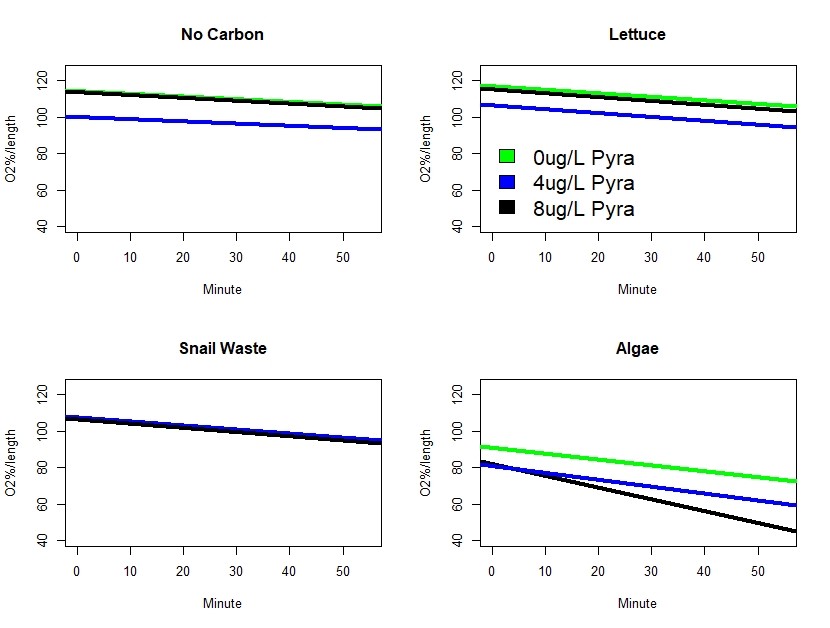
Finally, we attempted to determine and energetic signature of the above phenomenon by exposing daphnia to pyraclostrobin under the different carbon treatments (Figure 4). We hypothesized that strongest signal of an effect would occur in daphnia fed algae since this was the only energetically usable carbon form in the above studies. Although more data are needed to fully test the above hypothesis, algae-fed daphnia was the only treatment that showed a signal of the pyraclostrobin exposure. Specifically, at higher pyraclostrobin concentrations, there was an increase in oxygen consumption suggesting an increase in metabolic rate (see Algae figure above noting the slope of the black line). One hypothesis is that the increase in metabolic rate facilitated detoxification or elimination of pyraclostrobin thereby reducing toxicity. Collectively, these are interesting and promising results that point to both the importance of the carbon source and that the energetic environment can play a significant role in the manifestation of toxicity. In future research, my lab hopes to better define the interplay of these factors.
Specific Aim 2: Establish bioenergetics-based responses, life-history effects and population-level effects of individual study species exposed to PFOS and pyraclostrobin using a Life Table Response Experiment study design (full life cycle exposure).
Endpoints: Same as Specific Aim 1 plus population-level endpoints (population growth rate, estimated abundance, stable stage distribution)
Models: Individual-based and cohort-based population models
Summary Specific Aim 2 Progress: Full life-cycle, cohort-based study designs for Daphnia have now become a staple of our laboratory studies. This design involves initiating a “laboratory population” with a few individuals and allowing them to grow and reproduce with no culling for at least 40 days. This yields a very predictable population cycle of growth, peak, decline and stable phases. We have used this design to test toxicity of PFOS, NaCl and pyraclostrobin to Daphnia magna. In our most recent work, MS student Amanda Isabella has explored the interactive effects of chronic NaCl exposure with a pyraclostrobin pulse exposure occurring during the peak phase of the population cycle of D. magna.
Complex Chemical Mixtures – Impacts on Populations
Amanda Isabella, a second year M.S. student is conducting research to explore the impacts of chemical mixtures in complex ecological scenarios. Specifically, Amanda is interested in the effects of chemical mixtures and the role that non-consumptive predators play in modifying population dynamics and response to toxicants.
In her first major experiment, Amanda completed a population-level study in which laboratory populations of Daphnia magna (DM) were exposed to NaCl under chronic conditions and then received a pulse of pyraclostrobin at the peak of their population cycle. This scenario reflects one common to the Mid-Atlantic which is seeing increases in NaCl concentrations in ponds, lakes and streams largely from road de-icing salts. As mentioned in previous reports, the DM population studies that we now routinely employ include important intra-species interactions and dynamics and so, in that sense, are somewhat more ecologically realistic. The basic design ustilizes 1 liter glass jars with water changes three days per week. After 80-90% of the water has been siphoned, pictures are taken and DM enumerated and measured. These studies then yield a tremendous amount of data regarding abundance, size distribution and size-specific mortality.
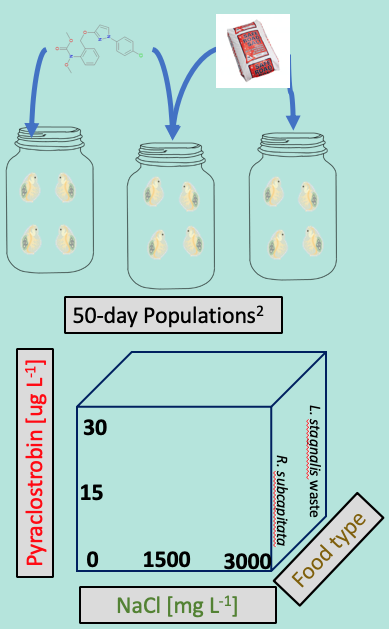
Figure 5. Experimental design of population exposure to determine interactive effects of NaCl and pyraclostrobin under more ecologically relevant conditions.
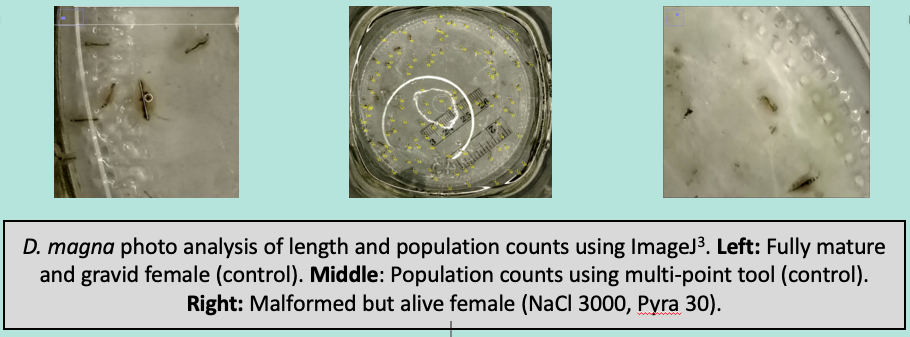
Figure 6 Obtaining data from DM population study using images captured during water changes.
As in other population studies with DM, the “typical” laboratory population dynamics were observed whereby there was sharp increase in abundance to a peak followed by a decline and then a stabilization of population size. There were effects of both NaCl and the pulse of pyraclostrobin (Fig. 7). In looking at DM abundance through time (Fig. 7) there is an overall depression of population size with increasing NaCl. There was then a sharp decline in population size around day 24 when the pyraclostrobin pulse occurred. Most interesting perhaps is the response of DM to the pyraclostrobin pulse across the different NaCl treatments. At 0 and 1500 mg/L NaCl, the highest pulse of pyraclostrobin (30 ug/L) showed the greatest decline followed by the largest compensation such that these populations had the highest abundance around day 40. For the 0 mg/L NaCl and high pyraclostrobin pulse, this population had the highest abundance for the remainder of the study. Even though the high pulse of pyraclostrobin clearly caused a shard and large decline, these treatments then recovered and exceeded abundance of the controls and lower pyraclostrobin pulse populations.Viewing the data as a percentage change from the control (Fig. 8), it is easier to see both the effects of NaCl (going from top to bottom figure) as well as the responses to the pyraclostrobin pulses. In all treatments of NaCl, the high pulse pyraclostrobin treatments had a compensatory response that was greater than the low (15 ug/L) and no (0 mg/L) pyraclostrobin pulses. These data are continuing to be analyzed but the large compensatory response in the high-pyraclostrobin pulse treatments was somewhat surprising. However, it seems reasonable to hypothesize that once abundance in these treatments dropped to the lowest level, remaining animals then had high resource levels and were able to capitalize and grow/reproduce at a higher rate. In essence, the higher pyraclostrobin pulse caused a release from density dependent effects on DM that were manifesting even at relatively low abundance levels. Although these data seem to reflect that DM exposed to high pulses of pyraclostrobin actually did better compared to low and no pulses, whether any lingering effects of pyraclostrobin remained is uncertain at this time.
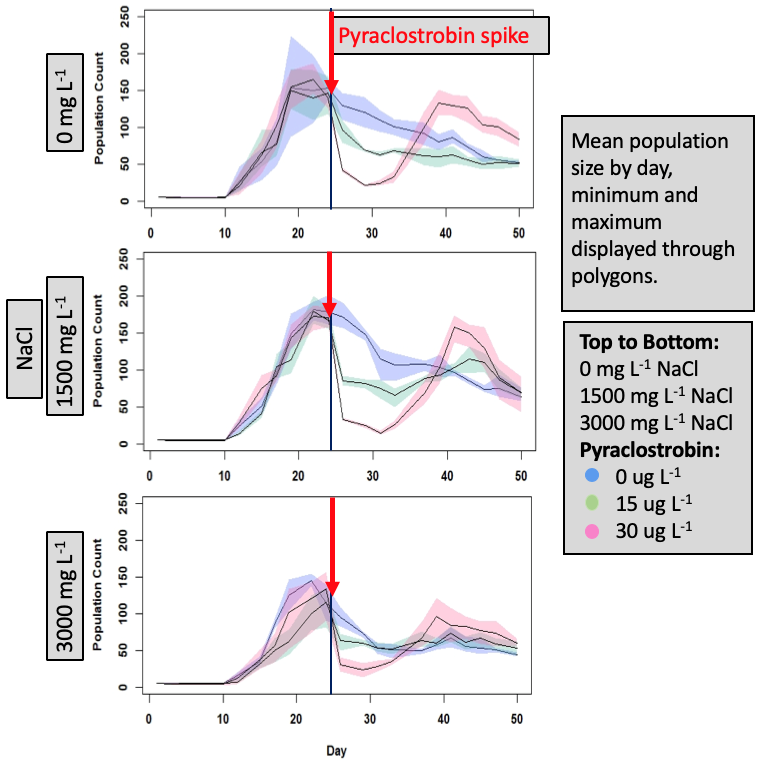
Figure 7. Daphnia magna abundance through time exposed to chronic, constant levels of NaCl with a pulse of pyraclostrobin occurring during the peak phase.
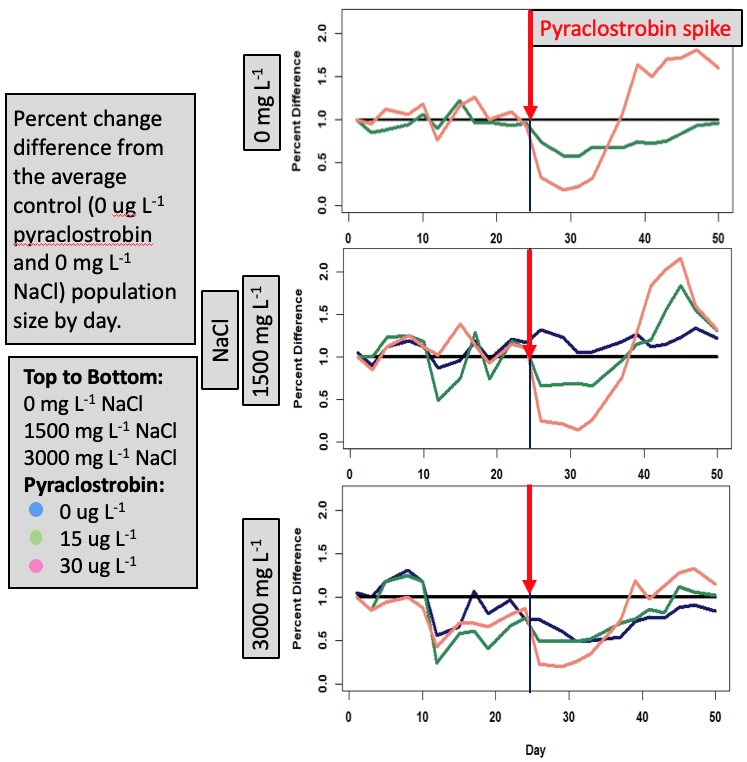
Figure 8. Daphnia magna percent different abundance compared to control through time exposed to chronic, constant levels of NaCl with a pulse of pyraclostrobin occurring during the peak phase.
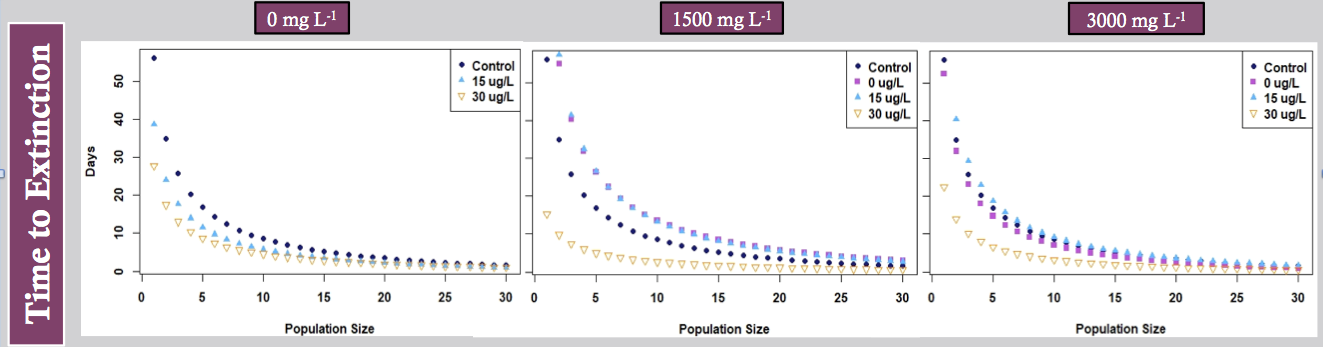
Figure 9. Time to extinction in Daphni magna laboratory populations at different concentrations of NaCl (different panels) and pyrclostrobin (curves within each panel).
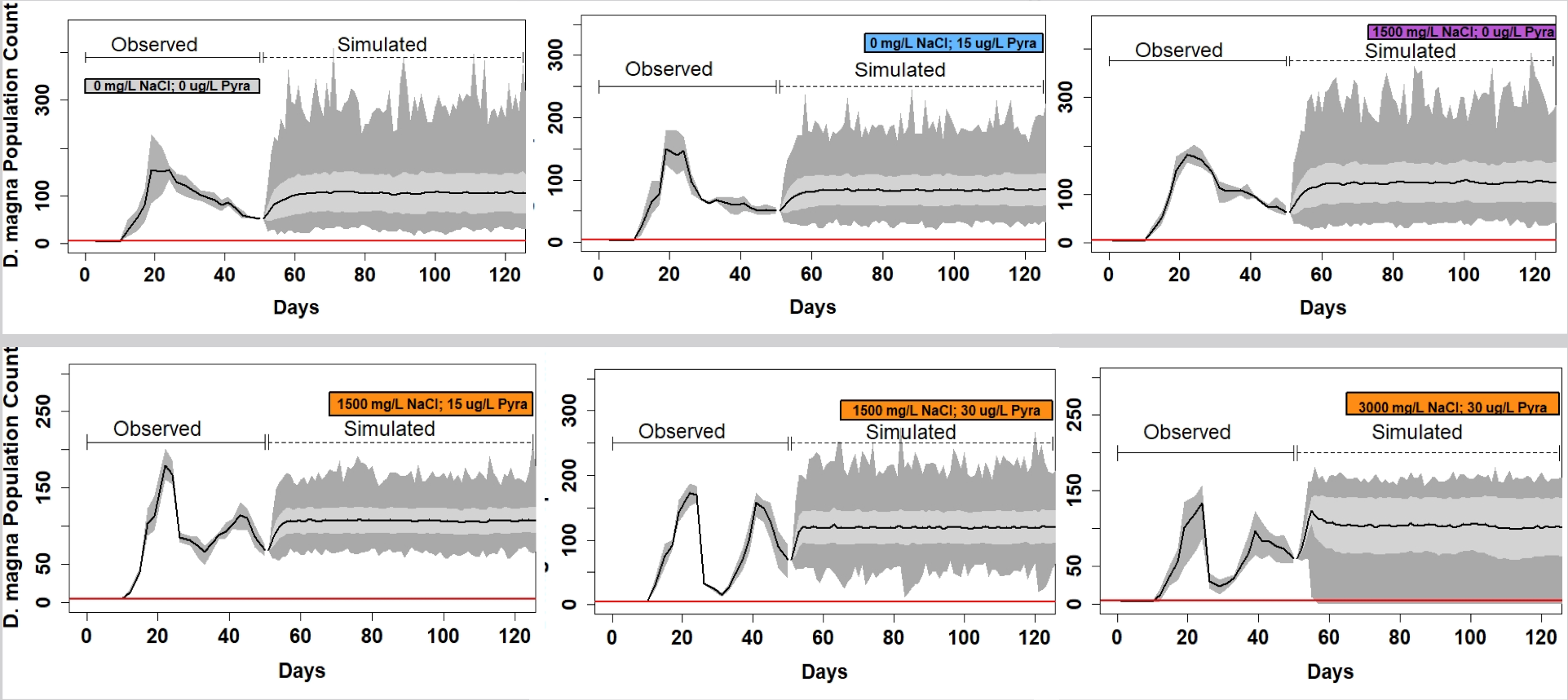
Figure 10. Simulations of a simple population model for the different treatments of NaCl and pyraclostrobin. Note the higher relative extinction risk in Daphnia exposed to the highest concentrations of NaCl and pyraclostrobin.
We developed a simple cohort-based population model to estimate population-level impacts of both NaCl and pyraclostrobin and the results are presented in Figures 9 and 10. Figure 9 expresses time to extinction based on different threshold population sizes (x-axis) or what is commonly termed quasi-extinction. This is implemented because at low population sizes, proceses such as demographic stochasticity can accelerate populations toward extinction. In all cases, the shortest time to extinction occurs in daphnia exposed to the highest pyraclostrobin concentrations. There does appear to be an effect of NaCl but it is not pronounced although the combination of the highest NaCl and pyraclostrobin concentrations do result in the highest relative extinction risk. We had previously developed a dynamic energy budget – individual based model (DEB-IBM) for daphnia that was spatially explicit and included crowding as a factor important to daphnia population dynamics. The model produces output that closely matches our experimental population data provided we have resource-specific toxicity values. The model has been applied to data generated by Tim Woo involving the effects of resource environment on toxicity of pyraclostrobin and will be used by Amanda Isabella to predict effects of two chemicals and non-consumptive predator effects in D. magna.
Specific Aim 3: Establish bioenergetics-based responses, life history effects and population-level effects of individual study species exposed to PFOS and pyraclostrobin in addition to the presence of conspecific competitors and predators.
Endpoints: Same as Specific Aim 1 plus modified life history based on species interactions (competition coefficients; predator-prey coefficients)
Models: Density dependent, resource based population models; Modified population models that include mortality from predation
Summary Specific Aim 3 Progress: We have completed studies in which we have combined L. stagnalis and D. magna and have been conducting studies over the last year that include predators of D. magna. In this scenario with L. stagnalis, the nature of the interaction is energetic facilitation to D. magna via L. stagnalis. Specifically, L. stagnalis “liberates” sources of energy (lettuce) making it available to D. magna. This is a relatively under studied form of species interaction but one that is likely common and potentially important. Also, our population study design for D. magna includes density-dependent effects and hence, intra-species competition. Our DEB-IBM also captures these dynamics well and illustrates one important attribute of IBMs – properties and dynamics of populations can emerge from the interactions and behaviors of individuals. To support the D. magna + L. stagnalis studies, we have developed a model that links DEB models for daphnia and snails and yields excellent agreement with study results (described previously). The model is still undergoing calibration and testing, but we are approaching what we think is a near final version. We have continued studies to explore the nonconsumptive effects of predation by fathead minnows on D. magna and have found very strong effects that have reduced the survival of Daphnia when exposed to non-consumptive predator cues although, interestingly, these effects appear to be ameliorated under some circumstances. Currently, Amanda Isabella is in the final stages of a larger laboratory population study in which daphnia are exposed to all combinations of elevated levels of Chloride (NaCl), a pulse of pyraclostrobin and predator kairomone water. This likely represents one of the last studies to be completed under the current proposal. An important and relatively untested ecological element in toxicity testing is predation. Predators exert direct (consumptive) and indirect (non-consumptive) effects on prey species. While consumptive effects are easy to visualize and quantify, indirect effects are equally or more impactful, but often harder to quantify. We are seeking a better understanding of how predators can modify the expression of toxicity when they exert indirect effects. Our study design originally involved fathead minnows (Pimephales promelas) caged within a larger tank that houses a DM population. Fish were fed 7 DM per day but do not have access to the DM in the rest of the tank. We have since changed the design to remove confounding effects associated with the caged fish.We first conducted a pilot study to evaluate the experimental design and to obtain a sense of how stressful these indirect predatory stressors were. To our surprise, the predator treatments showed high effects on DM survival (Fig. 11). We hypothesized that predators would negatively affect growth and reproduction but were surprised to find strong effects on survival. Clearly, nonconsumptive effects can have pronounced impacts on prey populations. Note that all water quality parameters (DO, conductivity, pH) were the same between predator and non-predator treatments.
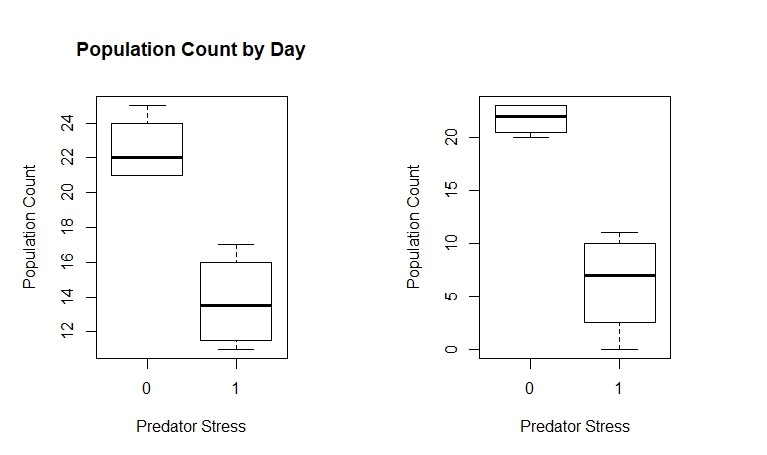
Figure 11. Effects of nonconsumptive predator (Pimephales promelas) cues on abundance of DM. There were strong and clear negative effects of the presence of predators on survival of DM. All treatments started with 25 DM individuals.
Future Activities:
The final daphnia population study will be completed March 6. The final report with therefore include a presentation and discussion of the results from this study as well as a previously conducted series of pilot studies.
Journal Articles on this Report : 2 Displayed | Download in RIS Format
| Other project views: | All 26 publications | 9 publications in selected types | All 9 journal articles |
|---|
| Type | Citation | ||
|---|---|---|---|
|
|
Reategui-Zirena EG, Fidder BN, Olson AD, Dawson DE, Bilbo TR, Salice CJ. Transgenerational endpoints provide increased sensitivity and insight into multigenerational responses of Lymnaea stagnalis exposed to cadmium. Environmental Pollution 2017;224:572-580. |
R835800 (2016) R835800 (2017) R835800 (2018) R835800 (2019) |
Exit Exit Exit |
|
|
Rohr JR, Salice CJ, Nisbet RM. The pros and cons of ecological risk assessment based on data from different levels of biological organization. Critical Reviews in Toxicology 2016;46(9):756-784. |
R835800 (2016) R835800 (2017) R835800 (2018) R835800 (2019) R835188 (Final) R835797 (2015) R835797 (2016) |
Exit Exit |
Progress and Final Reports:
Original AbstractThe perspectives, information and conclusions conveyed in research project abstracts, progress reports, final reports, journal abstracts and journal publications convey the viewpoints of the principal investigator and may not represent the views and policies of ORD and EPA. Conclusions drawn by the principal investigators have not been reviewed by the Agency.